The Effect of Frozen Storage on the Survival of Probiotic Microorganisms
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Old Fashioned Ricky Syrup, and Milk, 6.49 and Orange Juice, 7.49 Sparkling Soda Water, 4.99 Regular 3.49 Jumbo 4.59
Our Thick Frappe Tropical Fruit Frappe Your favorite syrup and three heaping scoops of A thick frappe blended with a fresh banana, 1. Select a Base: 2. Select a Size: ice cream blended with milk, 6.49 coconut pineapple ice cream, • Low-Fat • Single Portion 8 oz. / 5.29* and crushed pineapple fruit, 7.49 Extra Thick Frappe • Sugar-Free & Fat-Free • Double Portion 2 x 8 oz./ 8.29* Blended with 5 scoops of ice cream, 7.49 Oreo® Crunch Frappe • Non-Dairy *Includes 1 Mixing Flavor per 8 oz. portion Giant Thick Frappe Blended with Oreo® Ice Cream, syrup, ® Like our thick frappe but twice the size and and topped with crushed Oreos , 7.49 Served in a topped with whipped cream, 9.49 “PRU” Center Frappe Waffle Cone Chocolate Malted Frappe Your favorite syrup, 12 heaping scoops of ice 3. Select a Mixing Flavor: add 99¢ Our thick frappe blended with lots of malt cream blended with country fresh milk, topped • Vanilla* • Banana • Apple Pie • Butterfinger® with whipped cream -- over 2 quarts large! 17.99 and chocolate syrup, 7.49 • Chocolate* • Blueberry • Brownie • Chocolate Chip Cookie Dough • Coffee* • Chocolate Chips Sherbet Freeze • Lemon • Carrot Cake Real Banana Frappe A refreshing drink! Three heaping scoops • Espresso* • Heath Bar® A thick frappe blended with a fresh banana, • Maraschino Cherry • Chocolate Chip Cookie ® of tangy sherbet blended with soda. • Mocha* • M & M’s syrup and banana ice cream, 7.49 ® Your choice of Orange, Raspberry, or Lemon • Butterscotch • Orange • Ginger Snap Cookie • Mounds Real Strawberry Banana Frappe (Lime -

Copyrighted Material
Contents List of contributors xi Preface to the second edition xiii Preface to the first edition xv PART I BASIC BACKGROUND 1 1 History and consumption trends 3 Ramesh C. Chandan 1.1 Overview of the world dairy industry 3 1.2 Milk production in the USA 3 1.3 Production of dairy foods in the USA 4 1.4 Fermented/cultured dairy products 7 1.5 Occurrence and consumption of fermented milks in various regions 8 1.6 Major commercial fermented milks 11 1.7 Scandinavian fermented milks 15 1.8 Russian and Eastern European fermented milks 16 1.9 Middle Eastern fermented milks 16 1.10 South Asian fermented milks 17 References and further reading 19 2 Milk composition, physical and processing characteristics 21 Guillaume Brisson and Harjinder Singh 2.1 Introduction 21 2.2 Milk composition 21 2.3 Milk components and processing characteristics 23 2.4 Physicochemical properties of milk 37 References and further reading 44 3 RegulatoryCOPYRIGHTED requirements for milk production, MATERIAL transportation and processing 49 Cary P. Frye 3.1 Introduction 49 3.2 From farm to factory 49 3.3 History of milk safety 50 3.4 United States Public Health Grade “A” Milk Safety Program 50 3.5 Inspection of milk safety 51 3.6 Farm requirements 52 3.7 Milk transportation 53 00001668837.INDD001668837.INDD v 111/12/20121/12/2012 110:07:200:07:20 PPMM vi Contents 3.8 Processing plant 54 3.9 Hazard Analysis and Critical Control Point System 58 3.10 Standards and regulations 60 3.11 Milk pricing—US Federal Milk Marketing Orders 65 Glossary 68 References and further reading 68 4 Regulations for product standards and labeling 71 Cary P. -

Probiotic Recipes
LACTOSPORE ® Discover Functional Food Formulas LactoSpore® is a safe clinically tested probiotic that helps to maintain a healthy balance of microflora in the gastrointestinal tract, supporting general health and well being. Unlike many other probiotic cultures that do not survive processing and storage conditions, LactoSpore® being in naturally encapsulated spore form remains viable on processing and storage, and optimally delivers a live culture upon consumption. It beneficially impacts the microenvironment of the gastrointestinal tract by producing the biologically preferred L(+) form of lactic acid, and supporting a healthy balance of natural microflora; helping the body's natural defenses to effectively combat stress and imbalances. © 2015 Sabinsa !1 ProbiOaties Cookies, from BiteMarket, California, USA ProbiOaties Made in a bakery in sunny Southern California, BiteMarket exclusively sells these LactoSpore® containing cookies at their stores Use about 1/4 teaspoon of LactoSpore® 15B grade, for 10 to 12 small cookies that when baked are 2 inches in diameter. Bake them for approximately 10 minutes at 350° F (180°C). © 2015 Sabinsa !2 “Using LactoSpore in my cookies is a great way to incorporate probiotics into my life and the fact that they are shelf stable means that I can take them anywhere!” - BRIDGET REILLY, GLUTEN FREE GOODNESS FROM BRIDGET'S KITCHEN, OWNER ProbiOaties ProbiOaties Chocolate Chip Raisin Nutrition Nutrition Facts Facts © 2015 Sabinsa !3 Tutti Frutti’s Frozen Yogurt, available worldwide, sold to millions of delighted -
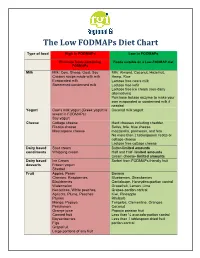
The Low Fodmaps Diet Chart
The Low FODMAPs Diet Chart Type of food High in FODMAPs Low in FODMAPs Eliminate foods containing Foods suitable on a Low-FODMAP diet FODMAPs Milk Milk: Cow, Sheep, Goat, Soy Milk: Almond, Coconut, Hazelnut, Creamy soups made with milk Hemp, Rice Evaporated milk Lactose free cow’s milk Sweetened condensed milk Lactose free kefir Lactose free ice cream (non-dairy alternatives) Purchase lactase enzyme to make your own evaporated or condensed milk if needed Yogurt Cow’s milk yogurt (Greek yogurt is Coconut milk yogurt lowest in FODMAPs) Soy yogurt Cheese Cottage cheese Hard cheeses including cheddar, Ricotta cheese Swiss, brie, blue cheese, Mascarpone cheese mozzarella, parmesan, and feta No more than 2 tablespoons ricotta or cottage cheese Lactose free cottage cheese Dairy based Sour cream Butter-limited amounts condiments Whipping cream Half and Half- limited amounts Cream cheese- limited amounts Dairy based Ice Cream Sorbet from FODMAPs friendly fruit desserts Frozen yogurt Sherbet Fruit Apples, Pears Banana Cherries, Raspberries, Blueberries, Strawberries Blackberries Cantaloupe, Honeydew-portion control Watermelon Grapefruit, Lemon, Lime Nectarines, White peaches, Grapes-portion control Apricots, Plums, Peaches Kiwi, Pineapple Prunes Rhubarb Mango, Papaya Tangelos, Clementine, Oranges Persimmon Coconut Orange juice Papaya passion fruit Canned fruit Less than ¼ avocado-portion control Boysenberries Less than 1 tablespoon dried fruit Figs portion control Grapefruit Large portions of any fruit Limit consumption to one low FODMAPs fruit -

Frozentreats Mindovermunch.Pdf
COPYRIGHT Alyssia Sheikh Frozen Treats: 40 Healthier Recipes! © 2018, Alyssia Sheikh Mind Over Munch® [email protected] ALL RIGHTS RESERVED. This eBook contains material protected under International and Federal Copyright Laws and Treaties. Any unauthorized reprint or use of this material is prohibited. No part of this eBook may be reproduced or transmi!ed in any form or by any means, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system without express wri!en permission from the author. www.mindovermunch.com All photos are property of Mind Over Munch®. DISCLAIMER: All content presented in this eBook is for informational purposes only. These statements have not been evaluated by the Food and Drug Administration. This product is not intended to diagnose, treat, cure or prevent any disease, nor is it intended to be a substitute for medical treatment. Please seek the advice of your healthcare professional for your specific health concerns. TABLE OF CONTENTS 1. Copyright 2. Master Table of Contents 3. Welcome Le!er 4. Legends & Notes POPSICLES ICE CREAM, SORBET, 5. Matcha Milk Popsicles FRO-YO & MILKSHAKES 6. Watermelon Green Tea Pops 25. Cherry Acai Ice Cream 7. Coconut Chai Pops 26. Avocado Ice Cream 8. Fizzy Raspberry Lemonade Pops 27. Banana Ice Cream 9. Caffeinated Chunky Monkey Pops 28. Raw Vegan Ice Cream Sandwiches 10. Strawberry Basil Limeade Slushy Pops 29. Cashew Vanilla Bean Ice Cream 11. Key Lime (Pie) Popsicles 30 Coconut Vanilla Ice Cream 12. Berry Oatmeal Breakfast Popsicles 31. Blueberry Sorbet 13. Strawberry Banana Pops 32. Strawberry Basil Sorbet 14. -

(Artocarpus Heterophyllus) Incorporated Frozen Yoghurt
International Journal of Scientific and Research Publications, Volume 9, Issue 6, June 2019 627 ISSN 2250-3153 Evaluation of Nutritional, Physico-chemical and Sensory Properties of Jackfruit (Artocarpus heterophyllus) Incorporated Frozen Yoghurt T.M.P.M. Dissanayaka, K.H.I. Gimhani, W.A.H. Champa Department of Animal and Food Sciences, Faculty of Agriculture, Rajarata University of Sri Lanka, Puliyankulama, Anuradhapura, Sri Lanka. DOI: 10.29322/IJSRP.9.06.2019.p9091 http://dx.doi.org/10.29322/IJSRP.9.06.2019.p9091 Abstract- The study was conducted to develop frozen yoghurt, flavoured with jackfruit pulp. Sensory data revealed that, the sample incorporated with 15% jackfruit pulp had the best qualities. Protein content of all jackfruit pulp incorporated frozen yoghurts was significantly higher (p < 0.05) and their fat content was significantly lower (p < 0.05) compared to the control. Significantly higher (p < 0.05) overrun was observed in the frozen yoghurt containing 20% of jackfruit pulp compared to the control. Titratable acidity increased and pH decreased in all samples during the storage. Though total plate count of all jackfruit pulp incorporated frozen yoghurts increased upon storage, it was less than that of the control. Escherichia coli, yeast and mould were not detected for a period of 35 days at frozen storage. In conclusion, physicochemical, microbiological, and sensory properties of frozen yoghurt improved significantly by adding jackfruit pulp at rate of 15% (w/w). Index Terms- Frozen yoghurt, Jackfruit, Quality evaluation, Value addition I. INTRODUCTION Cow milk contains high uality proteins with a good balance of all essential amino acids and minerals, especially calcium and phosphorous than human milk (FAO, 2013). -

Frozen Yogurt
Shakes Smoothies Make it a Malt $1.00 All Fruit, Dairy-Free $1.00 Add a Boost $1.00 per boost Add a Boost $1.00 per boost Add Mix-In $1.00 Add Mix-In $1.00 Add Fresh Fruit $1.50 Add Fresh Fruit $1.50 vanilla $5.99 Acaí Brunch Bowl $9.99 Strawberry Banana $5.99 Vanilla Frozen Yogurt, Vanilla Ice Cream, A blend of acaí, blueberries, banana, and apple juice, topped with Apple Juice, Strawberries, Banana, Whipped Cream honey-almond granola, fresh strawberries, and a drizzle of honey. Strawberry Sorbet, Non-fat Frozen Yogurt Add a seasonal fresh fruit, topping, $ $ Chocolate 5.99 boost or extra granola for $1.00 each. Hawaiian Breeze 5.99 Chocolate Frozen Yogurt, Ghirardelli Chocolate Sauce, Passion Fruit-Mango Juice, Pineapple, Banana, Sweet Things Suggests: Vanilla Ice Cream, Whipped Cream Pineapple Sherbet, Non-fat Frozen Yogurt - Peanut Butter & Chocolate Chips - Seasonal Fresh Fruit, Almonds & Coconut Cookies & Cream $6.99 - Soy Protein or Whey Protein Mango Madness $5.99 Vanilla Frozen Yogurt, Oreos, Passion Fruit-Mango Juice, Mango, Banana, Vanilla Ice Cream, Whipped Cream Add a Boost for $1.00 Non-fat Frozen Yogurt Whey Protein • Soy Protein • Immunity • Greens • Energy Strawberry Sensation $6.99 Strawberry Sunrise $5.99 Vanilla Frozen Yogurt, Strawberries, Strawberry Sorbet, Apple Juice, Strawberries, Pineapple, Strawberry Sorbet, Vanilla Ice Cream, Whipped Cream Greek Yogurt Parfait $7.99 Pineapple Sherbet, Non-fat Frozen Yogurt Protein-packed non-fat Greek yogurt, honey-almond granola, your Peanut Butter Power $6.99 choice of two seasonal fresh fruits or toppings, and a drizzle of honey. -

“Frozen Yogurt Manufacturing”
FROZEN YOGURT MANUFACTURING Item Type text; Electronic Thesis Authors COOK, KATHRYN LOUISE KRAMER; Moline, Dena; Morehead, Yirla; Ormsby, Stefka Publisher The University of Arizona. Rights Copyright © is held by the author. Digital access to this material is made possible by the University Libraries, University of Arizona. Further transmission, reproduction or presentation (such as public display or performance) of protected items is prohibited except with permission of the author. Download date 30/09/2021 18:37:15 Item License http://rightsstatements.org/vocab/InC/1.0/ Link to Item http://hdl.handle.net/10150/190435 Frozen Yogurt Manufacturing Course: ChEE 443: Plant Design Submitted: May 2, 2008 _______________________________ _______________________________ Kathryn Cook, Senior Design Engineer Dena Moline, Senior Design Engineer _______________________________ _______________________________ Yirla Morehead, Senior Design Engineer Stefka Ormsby, Senior Design Engineer Executive Summary The goal of this project was to design a manufacturing facility to produce 7.4 million containers of vanilla frozen yogurt annually with each container holding 1.75 quarts. Pennsylvania was chosen as the building site because of the demand for frozen yogurt in the region. In today’s market there are many reasons to market more low-fat and more healthful foods based on recent diet and health trends. In 2006, the market for frozen yogurt in the United States was $200 million, and 30% of this market is concentrated in the northeastern US alone (Parker, 2005). While Yogurina will supply a small percentage of that, 5%, we feel that the region has a strong demand for the low-fat frozen yogurt product we are offering. Frozen yogurt has several key ingredients, starting with ultrafiltered skim milk. -

Download the Guide
ONTARIO 2021 DAIRY GUIDE ONTARIO DAIRY GUIDE 1 Whether crafted by hand or at scale, Ontario dairy products are all inspired by a single delicious, sustainable and local ingredient: EVERYDAY MILK… DID YOU KNOW? • As a result of continuous investments in sustainability LOCAL MILK WHOLE MILK and production practices, 65% fewer cows produce Barista milk! Whole (3.25% milk fat) milk is lighter than cream but the same volume of milk needed to nourish all of Producing the fresh, local milk that elevates an incredible range of high-quality adds a luxurious smoothness to coffee beverages and dishes. Canada as compared with 50 years ago. Ontario dairy products is a tradition based on the values that have shaped dairy LOCAL MILK LOCAL farming for generations. While the know-how and craft have been preserved, the PARTLY-SKIMMED & SKIMMED MILK • It takes the same amount of land resources quality and potential of the ingredient has grown and evolved, making Ontario one Partly-skimmed (1-2% milk fat) milk perfectly versatile, used in (about 1.7 square metres) to produce either a litre of the most unique dairy regions in the country and around the world. baking, cooking or enjoyed on its own. Skimmed milk (0.1% milk fat) of milk or a loaf of bread. is nearly fat-free and extremely refreshing. • A single serving of milk contains more absorbable BUTTERMILK calcium than any other natural food. LOCAL BY NATURE. Perfected. Pasteurization eliminates bacteria and The inimitable tanginess of buttermilk is the result of a specialized Milk is a hyper local food, travelling from the farm to grocery homogenization ensures a smooth and consistent milk bacterial culture added to regular milk. -

Winners and Finalists of the 2017 World Dairy Innovation Awards!
Winners and finalists of the 2017 World Dairy Innovation Awards! Last night the annual World Dairy Innovation Awards, in association with FoodBev Media, took place at the 11th Global Dairy Congress, in Dublin. The judging panel considered entries from over 20 countries in 19 categories. “This is the eleventh annual World Dairy Innovation Awards, and once again we are fascinated by the level of innovation that is shown within the entries. We see new flavours, new concepts, clever packaging, state-of-the- art manufacturing technologies and new ground gained towards a more responsible and sustainable dairy industry,” said FoodBev Media marketing director Isabel Sturgess. “World Dairy Innovation Awards really showcase some of the most ambitious new products and developments of the year, from a wide range of companies all over the world, both large and small. Keep an eye on the latest entries to spot the upcoming trends.” A full list of finalists and winners in each category follows below. Best Ice Cream or Frozen Yogurt Winner Yomega3 Greek Style Frozen Yogurt – Ice Cream Treats Finalists Baileys Salted Caramel Ice Cream – Bulla Dairy Foods Baileys Chocolat Luxe Ice Cream – Bulla Dairy Foods Yasso’s Dessert – Yasso Yo2 – Dairy Farmers of America Best Yogurt Winner DεLTα Mini Meals – Delta Foods S. A. Finalists Kerrygold Rhubarb Yogurt – Ornua Natural Durian Stirred Yogurt - TH true YOGURT Kids Yogurt - Tamar Valley Dairy Bulgarian Rose Yogurt – Balkan Treasure Icelandic skyr – MS Iceland Dairies Best Cheese Winner Hokkaido Tokachi Rich 3-Way Cheese Slices – Meiji Co. Ltd Finalists Hay Flower Rebel - Käserebellen GmbH Cracker Cuts - Dutch Farms Inc. -

ROSAUERS Maxwell House Premium Roast International Beverage Mix Bulk Coffee ROSAUERS ROSAUERS 4-Oz
AD STARTS JANUARY 29! Full Circle Festival 100% Market Tostitos Pure Coconut Organic Coffee Water Tortilla Chips • 12-oz. Ground 17.5-oz. 10-oz. to 13-oz. • 12-ct. Single Serve Cups Original or With Pulp Selected Varieties Selected Varieties $299 $589 PRICES EFFECTIVE JAN. 29 THRU FEB. 25, 2020 We Accept... EBT Quest Cards/W.I.C. 1 - ALL Keebler Town House or Club Crackers 9-oz. to 13.8-oz. Selected Varieties $299 Not Available Oroweat in Libby, MT Bread 24-oz. Selected Varieties $199 Oroweat Not Available English Muf�ins in Libby, MT 6-ct. Selected Varieties Franz $ 59 Bread 1 22.5-oz. or 24-oz. Moonlight Mixes Moonlight Mixes Wicked Mix Selected Varieties Wicked Minis 7-oz. Selected Varieties Oyster Crackers $ 99 6-oz. 4 $ 29 Selected Varieties 2|$5 2 - ALL 2 EFFECTIVE DATES 01/29/20 - 02/25/20 Nabisco Ritz Crackers 7.5-oz. to 13.7-oz. Selected Varieties $279 Nabisco Snack Crackers 6.5-oz. to 16-oz. Family Size! Selected Varieties $329 Nabisco Premium Saltine Crackers 24-oz. Family Size! $ 29 Nabisco 3 Oreo Cookies Nabisco 7.9-oz. to 15.35-oz. Ritz Snacks Selected Varieties 7.1-oz. or 8.1-oz. Selected Varieties 2 $ $279 | EFFECTIVE DATES 01/29/20 - 02/25/20 5 3 - ALL Earthwise Organic Juice Beverages 52-oz., Chilled Selected Varieties 2|$4 Donald Duck Orange Juice 59-oz., Chilled Selected Varieties $ 49 Florida's Natural Apple or 2 Orange Juice 14-oz., Chilled Langers $129 100% Juice 64-oz. Florida's Selected Varieties Natural $ 49 Fruit Juice 2 52-oz. -

Crediting Foods in the CACFP
Crediting Foods in the CACFP All meals claimed for reimbursement must meet the CACFP Meal Pattern requirements. This includes serving foods that are creditable, meeting the meal pattern components and minimum serving sizes. Refer to Guidance Memorandum 12 for details on the meal pattern. Milk Non-Creditable Milk (Not an all-inclusive list) Required at breakfast, lunch, and supper • 2% (reduced fat) milk Fluid milk can be served as a beverage, used on cereal, or used in part for both • Almond Milk Creditable milks: Pasteurized cow’s and goat’s milk, lactose-free and lactose-reduced milk, fat- • Cheese (creditable as meat alternate, not as milk) free or low-fat buttermilk, and acidified milk • Coconut Milk • Cream Cheese 1 year olds must be served unflavored whole milk • Cream Soup/Sauces 2-5 year olds must be served unflavored 1% or skim • Evaporated Milk 6-18 year olds must be served unflavored or flavored 1% or skim • Frozen Yogurt Fluid milk cannot be served to infants (less than 12 months of age—medical statement required) • Ice cream, Ice Milk Milk used in cooking (i.e. cooked cereals, custards, pudding, etc.) cannot be credited • Powdered Milk • Pudding Milk may not be served for snacks when juice is served as the other component • Raw Milk • Rice Milk Non-dairy milk substitutes that are nutritionally equivalent to fluid cow’s milk may be provided to • Reconstituted Dry Milk participants when supported by a written statement from a participant’s parent or legal guardian • Sour Cream Currently only some soy milk products are creditable. Refer to GM 12.