Palynology of an Extinct Barrier Island Wetland, Isle of Hope, GA
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Zephaniah Kingsley, Slavery, and the Politics of Race in the Atlantic World
Georgia State University ScholarWorks @ Georgia State University History Theses Department of History 2-10-2009 The Atlantic Mind: Zephaniah Kingsley, Slavery, and the Politics of Race in the Atlantic World Mark J. Fleszar Follow this and additional works at: https://scholarworks.gsu.edu/history_theses Recommended Citation Fleszar, Mark J., "The Atlantic Mind: Zephaniah Kingsley, Slavery, and the Politics of Race in the Atlantic World." Thesis, Georgia State University, 2009. https://scholarworks.gsu.edu/history_theses/33 This Thesis is brought to you for free and open access by the Department of History at ScholarWorks @ Georgia State University. It has been accepted for inclusion in History Theses by an authorized administrator of ScholarWorks @ Georgia State University. For more information, please contact [email protected]. THE ATLANTIC MIND: ZEPHANIAH KINGSLEY, SLAVERY, AND THE POLITICS OF RACE IN THE ATLANTIC WORLD by MARK J. FLESZAR Under the Direction of Dr. Jared Poley and Dr. H. Robert Baker ABSTRACT Enlightenment philosophers had long feared the effects of crisscrossing boundaries, both real and imagined. Such fears were based on what they considered a brutal ocean space frequented by protean shape-shifters with a dogma of ruthless exploitation and profit. This intellectual study outlines the formation and fragmentation of a fluctuating worldview as experienced through the circum-Atlantic life and travels of merchant, slaveowner, and slave trader Zephaniah Kingsley during the Era of Revolution. It argues that the process began from experiencing the costs of loyalty to the idea of the British Crown and was tempered by the pervasiveness of violence, mobility, anxiety, and adaptation found in the booming Atlantic markets of the Caribbean during the Haitian Revolution. -

Okefenokee Swamp and St. Marys River Named Among America's
Okefenokee Swamp and St. Marys River named Among America’s Most Endangered Rivers of 2020 Mining threatens, fish and wildlife habitat; wetlands; water quality and flow Contact: Ben Emanuel, American Rivers, 706-340-8868 Christian Hunt, Defenders of Wildlife 828-417-0862 Rena Ann Peck, Georgia River Network, 404-395-6250 Alice Miller Keyes, One Hundred Miles, 912-230-6494 Alex Kearns, St. Marys EarthKeepers, 912-322-7367 Washington, D.C. –American Rivers today named the Okefenokee Swamp and St. Marys River among America’s Most Endangered Rivers®, citing the threat titanium mining would pose to the waterways’ clean water, wetlands and wildlife habitat. American Rivers and its partners called on the U.S. Army Corps of Engineers and other permitting agencies to deny any proposals that risk the long-term protection of the Okefenokee Swamp and St. Marys River. “America’s Most Endangered Rivers is a call to action,” said Ben Emanuel, Atlanta- based Clean Water Supply Director with American Rivers. “Some places are simply too precious to allow risky mining operations, and the edge of the unique Okefenokee Swamp is one. The Army Corps of Engineers must deny the permit to save this national treasure.” The annual America’s Most Endangered Rivers report is a list of rivers at a crossroads, where key decisions in the coming months will determine the rivers’ fates. Over the years, the report has helped spur many successes including the removal of outdated dams, the protection of rivers with Wild and Scenic designations, and the prevention of harmful development and pollution. Rena Ann Peck, Executive Director of Georgia River Network, explains "The Okefenokee Swamp is like the heart of the regional Floridan aquifer system in southeast Georgia and northeast Florida. -

Georgia Historical Society Educator Web Guide
Georgia Historical Society Educator Web Guide Guide to the educational resources available on the GHS website Theme driven guide to: Online exhibits Biographical Materials Primary sources Classroom activities Today in Georgia History Episodes New Georgia Encyclopedia Articles Archival Collections Historical Markers Updated: July 2014 Georgia Historical Society Educator Web Guide Table of Contents Pre-Colonial Native American Cultures 1 Early European Exploration 2-3 Colonial Establishing the Colony 3-4 Trustee Georgia 5-6 Royal Georgia 7-8 Revolutionary Georgia and the American Revolution 8-10 Early Republic 10-12 Expansion and Conflict in Georgia Creek and Cherokee Removal 12-13 Technology, Agriculture, & Expansion of Slavery 14-15 Civil War, Reconstruction, and the New South Secession 15-16 Civil War 17-19 Reconstruction 19-21 New South 21-23 Rise of Modern Georgia Great Depression and the New Deal 23-24 Culture, Society, and Politics 25-26 Global Conflict World War One 26-27 World War Two 27-28 Modern Georgia Modern Civil Rights Movement 28-30 Post-World War Two Georgia 31-32 Georgia Since 1970 33-34 Pre-Colonial Chapter by Chapter Primary Sources Chapter 2 The First Peoples of Georgia Pages from the rare book Etowah Papers: Exploration of the Etowah site in Georgia. Includes images of the site and artifacts found at the site. Native American Cultures Opening America’s Archives Primary Sources Set 1 (Early Georgia) SS8H1— The development of Native American cultures and the impact of European exploration and settlement on the Native American cultures in Georgia. Illustration based on French descriptions of Florida Na- tive Americans. -

Plant Succession on Burned Areas in Okefenokee Swamp Following the Fires of 1954 and 1955 EUGENE CYPERT Okefenokee National Wildlife Refuge U.S
Plant Succession on Burned Areas in Okefenokee Swamp Following the Fires of 1954 and 1955 EUGENE CYPERT Okefenokee National Wildlife Refuge U.S. Bureau of Sport Fisheries and 'Wildlife Waycross, GA 31501 INTRODUCTION IN 1954 and 1955, during an extreme drought, five major fires occurred in Okefenokee Swamp. These fires swept over approximately 318,000 acres of the swamp and 140,000 acres of the adjacent upland. In some areas in the swamp, the burning was severe enough to kill most of the timber and the understory vegetation and burn out pockets in the peat bed. Burns of this severity were usually small and spotty. Over most of the swamp, the burns were surface fires which generally killed most of the underbrush but rarely burned deep enough into the peat bed to kill the larger trees. In many places the swamp fires swept over lightly, burning surface duff and killing only the smaller underbrush. Some areas were missed entirely. On the upland adjacent to the swamp, the fires were very de structive, killing most of the pine timber on the 140,000 acres burned over. The destruction of pine forests on the upland and the severe 199 EUGENE CYPERT burns in the swamp caused considerable concern among conservation ists and neighboring land owners. It was believed desirable to learn something of the succession of vegetation on some of the more severely burned areas. Such knowl edge would add to an understanding of the ecology and history of the swamp and to an understanding of the relation that fires may have to swamp wildlife. -

From the Vaults, February 2017
FROM The VAULTS Newsletter of the Georgia Archives www.GeorgiaArchives.org Volume 2, No. 1 February 2017 The Georgia Archives Building State Historian and Archives Director Louise Hays thought that the Georgia Archives would be preparing to move to a new building on the Capitol Square with the State Library and perhaps the Supreme Court as soon as World War II was over. It took another twenty years, a new State Archivist (Mary Givens Bryan), and a new Secretary of State (Ben W. Fortson, Jr.) to complete the white marble structure on Capitol Avenue. The Archives had moved from the fourth floor of the Capitol in 1929 and 1930 to the twenty-room mansion donated by the heirs of furniture magnate Amos G. Rhodes at 1516 Peachtree Street NW. From the beginning it was obvious that a fireproof building designed as an archives would be preferable to Rhodes Hall. Governor Lamartine Griffin Hardman in 1931 proposed carving a giant records storage facility into the core of Stone Mountain. Continued on page 8 Volume 2, No. 1 FROM THE VAULTS Page 2 News From Friends of Georgia Archives Update from the President Welcome to Spring in Georgia and at the Georgia Archives! Officers and members of the Friends of Georgia Archives hope you will be able to join us for some of the activities at the Georgia Archives. Plus, the Archives is the perfect spot for researching your genealogy or state history and getting ready for family reunions! This newsletter is filled with information about activities at the Georgia Archives. Read through and mark your calendar now to attend some or all of them. -

A Vegetative Survey of Back-Barrier Islands Near Sapelo Island, Georgia
A VEGETATIVE SURVEY OF BACK-BARRIER ISLANDS NEAR SAPELO ISLAND, GEORGIA Gayle Albers1 and Merryl Alber2 AUTHORS: 1Graduate Student, Institute of Ecology and 2Associate Professor, Dept. of Marine Sciences, University of Georgia, Athens, GA 30602. REFERENCE: Proceedings of the 2003 Georgia Water Resources Conference, held April 23-24, 2003, at the University of Georgia. Kathryn J. Hatcher, editor, Institute of Ecology, The University of Georgia, Athens, Georgia. Abstract. This study was designed to examine the located in McIntosh County comprising 20% of the forest composition, structure and species richness of area (CMHAC, 2002). vegetation among undeveloped back-barrier islands Due to increased development pressure in coastal near Sapelo Island, Georgia. Known colloquially as Georgia, back-barrier islands have become a topic of “marsh hammocks,” back-barrier islands are debate. An administrative court suit resulted in the completely or partially encircled by estuarine salt appointment of Georgia’s coastal marsh hammock marsh. There are upwards of 1200 hammocks along advisory council (CMHAC) in February 2001 to assess the Georgia coast, comprising approximately 6900 ha. hammock development issues and define Georgia’s In the face of increased development pressure, the coastal marsh hammocks. The working definition cumulative impacts caused by small-scale construction adopted by the CMHAC was: of homes, roads, bridges, and septic fields may alter Back–barrier islands are all other1 islands between natural hydrologic and ecological processes. We the landward boundary of the barrier island surveyed vegetation on 11 undeveloped hammocks in complexes and the mainland. Natural back-barrier four size classes and found that overall species diversity islands are erosional remnants of pre-existing is low, but the diversity of vascular plants may increase upland, whereas man-made back barrier islands with island size. -

* This Is an Excerpt from Protected Animals of Georgia Published By
Common Name: BLACKBANDED SUNFISH Scientific Name: Enneacanthus chaetodon Other Commonly Used Names: none Previously Used Scientific Names: none Family: Centrarchidae Rarity Ranks: G4/S1 State Legal Status: Endangered Federal Legal Status: Not Listed Description: The blackbanded sunfish is a small, laterally compressed and deep-bodied species reaching a maximum total length of 100 mm (4 inches). There is a prominent notch separating the spinous and soft-rayed portions of the dorsal fin. It is distinctively marked with 5-6 black bars along the sides that extend from the dorsum to the venter. The first of these bars passes through the eye, and the third extends through the first three membranes of the spinous dorsal fin to the upper edge of the fin. No other sunfish has this barring pattern. The blackbanded sunfish is also very colorful with black vertical bars, olive-brown to variegated-brown on the dorsum and upper sides, and orange-copper marking the leading edge of the pelvic fins and the irises. Similar Species: The small body size and distinctive color pattern make it difficult to confuse the blackbanded sunfish with any other fish species in Georgia waters. It may superficially resemble the banded (Enneacanthus obesus) and bluespotted (E. gloriosus) sunfishes, which differ in having only a shallow notch separating the spinous and soft-rayed portions of the dorsal fin and lacking the prominent dark bar extending through the anterior dorsal fin membranes. Habitat: Blackbanded sunfish are restricted to shallow, low-velocity, non-turbid waters of lakes, ponds, rivers and streams. They are strongly associated with aquatic plants, which provide habitat for foraging and cover. -

Final General Management Plan, Wilderness Study, and Environmental Impact Statement June 2013
Fort Pulaski National Monument National Park Service Georgia U.S. Department of the Interior Final General Management Plan, Wilderness Study, and Environmental Impact Statement June 2013 Front cover photo credits, clockwise from top left:National Park Service; Tammy Herrell; David Libman; David Libman General Management Plan / Wilderness Study / Environmental Impact Statement Fort Pulaski National Monument Chatham County, Georgia SUMMARY President Calvin Coolidge established Fort Lighthouse and the other is known as Pulaski as a national monument by Daymark Island. Finally, in 1996, Congress proclamation on October 15, 1924, under the passed a law that removed the U.S. Army authority of section 2 of the Antiquities Act Corps of Engineers’ reserved right to deposit of 1906. The proclamation declared the dredge spoil on Cockspur Island. entire 20-acre area “comprising the site of the old fortifications which are clearly This General Management Plan / Wilderness defined by ditches and embankments” to be Study / Environmental Impact Statement a national monument. provides comprehensive guidance for perpetuating natural systems, preserving By act of Congress on June 26, 1936 (49 Stat. cultural resources, and providing 1979), the boundaries of Fort Pulaski opportunities for high-quality visitor National Monument were expanded to experiences at Fort Pulaski National include all lands on Cockspur Island, Monument. The purpose of the plan is to Georgia, then or formerly under the decide how the National Park Service can jurisdiction of the secretary of war. The best fulfill the monument’s purpose, legislation also authorized the Secretary of maintain its significance, and protect its the Interior to accept donated lands, resources unimpaired for the enjoyment of easements, and improvements on McQueens present and future generations. -

Case 2:16-Cv-00053-RSB-BWC Document 199 Filed 03/07/19 Page 1 of 100
Case 2:16-cv-00053-RSB-BWC Document 199 Filed 03/07/19 Page 1 of 100 IN THE UNITED STATES DISTRICT COURT FOR THE SOUTHERN DISTRICT OF GEORGIA BRUNSWICK DIVISION SARAH FRANCES DRAYTON; CAROLYN BANKS; MELVIN BANKS, SR.; CEASER BANKS; NANCY BANKS; LORIE BANKS on behalf of herself and the ESTATE OF MELVIN BANKS, JR.; MARION BANKS; ROBERTA BANKS; RICHARD BANKS; ELLEN BROWN; EARLENE DAVIS; ANDREA DIXON; DEBORAH DIXON; SAMUEL L. DIXON; DAN GARDNER; CHERYL GRANT; BOBBY GROVNER; CELIA GROVNER; DAVID GROVNER, SR., on behalf of himself and the ESTATE OF VERNELL GROVNER; DAVID GROVNER, JR.; IREGENE GROVNER, JR.; IREGENE GROVNER ,SR.; RALL GROVNER; ANGELA HALL; ANGELINA Case No. 2:16-cv-00053-DHB-RSB HALL; REGINALD HALL; BENJAMIN HALL; FLORENCE HALL; JOSEPH HALL; SECOND AMENDED MARGARET HALL on behalf of herself and COMPLAINT FOR DAMAGES the ESTATE OF CHARLES HALL; AND DECLARATORY AND VICTORIA HALL; ROSEMARY HARRIS; INJUNCTIVE RELIEF DENA MAY HARRISON on behalf of the ESTATE OF HAROLD HILLERY; JOHNNIE HILLERY; BRENDA JACKSON; JURY TRIAL DEMANDED JESSE JONES; TEMPERANCE JONES; SONNIE JONES; HARRY LEE JORDAN; DELORES HILLERY LEWIS; JOHNNY MATTHEWS; FRANCES MERCER; MARY DIXON PALMER; LISA MARIE SCOTT; ANDREA SPARROCK; DAVID SPARROCK; AARON WALKER; VERDIE WALKER; MARCIA HALL WELLS; STACEY WHITE; SYLVIA WILLIAMS; VALERIE WILLIAMS; HELP ORG, INC.; and RACCOON HOGG, CDC, Plaintiffs, v. MCINTOSH COUNTY, GEORGIA, by and 1 Case 2:16-cv-00053-RSB-BWC Document 199 Filed 03/07/19 Page 2 of 100 through its BOARD OF COMMISSIONERS; STATE OF GEORGIA; GOVERNOR NATHAN DEAL, in his official capacity; GEORGIA DEPARTMENT OF NATURAL RESOURCES; GEORGIA DEPARTMENT OF NATURAL RESOURCES COMMISSIONER MARK WILLIAMS, in his official capacity; GEORGIA DEPARTMENT OF COMMUNITY AFFAIRS; and MCINTOSH COUNTY SHERIFF STEPHEN JESSUP, in his official capacity, Defendants. -

Analysis of Shoreline Change for Jekyll and Sapelo Islands, Georgia
ANALYSIS OF SHORELINE CHANGE FOR JEKYLL AND SAPELO ISLANDS, GEORGIA WITH GIS TECHNIQUES by BEI TU Under the Direction of E. Lynn Usery ABSTRACT The objective of this research is to use geographic information systems (GIS) to quantify shoreline position change during 1954 and 1999 for Sapelo Island and Jekyll Island in Georgia, USA. Shorelines from multiple years were manually traced from digital raster graphics, aerial photographs, digital orthophoto quad quadrangles and a lidar image using ArcView GIS software, Version 3.3. This study showed that the northern ends of Jekyll and Sapelo islands are eroding, whereas the southern ends are accreting. Mann-Whitney test (two-tailed) indicated that the mean accretion rates and annual change rates (regardless of direction) of the two islands differed statistically during 1954-1974,1974-1993 and 1954-1999. The mean recession rates were significantly different during 1954-1974 and 1974-1993, but not so for 1954-1999. Human activity exerted a heavy influence on the shoreline change. Quantifying shoreline change provides useful data on the effects of engineering structures on erosion and is important for coastal zone management and planning. INDEX WORDS: GIS, shoreline change, Sapelo Island, Jekyll Island, lidar, coastal zone ANALYSIS OF SHORELINE CHANGE FOR JEKYLL AND SAPELO ISLANDS, GEORGIA WITH GIS TECHNIQUES by BEI TU B.E., Wuhan University, P. R. China, 2001 A Thesis Submitted to the Graduate Faculty of The University of Georgia in Partial Fulfillment of the Requirements for the Degree MASTER OF SCIENCE ATHENS, GEORGIA 2004 © 2004 Bei Tu All Rights Reserved ANALYSIS OF SHORELINE CHANGE FOR JEKYLL AND SAPELO ISLANDS, GEORGIA WITH GIS TECHNIQUES by BEI TU Major Professor: E. -
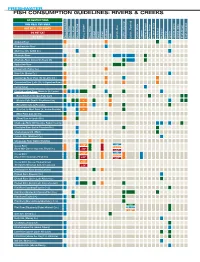
Fish Consumption Guidelines: Rivers & Creeks
FRESHWATER FISH CONSUMPTION GUIDELINES: RIVERS & CREEKS NO RESTRICTIONS ONE MEAL PER WEEK ONE MEAL PER MONTH DO NOT EAT NO DATA Bass, LargemouthBass, Other Bass, Shoal Bass, Spotted Bass, Striped Bass, White Bass, Bluegill Bowfin Buffalo Bullhead Carp Catfish, Blue Catfish, Channel Catfish,Flathead Catfish, White Crappie StripedMullet, Perch, Yellow Chain Pickerel, Redbreast Redhorse Redear Sucker Green Sunfish, Sunfish, Other Brown Trout, Rainbow Trout, Alapaha River Alapahoochee River Allatoona Crk. (Cobb Co.) Altamaha River Altamaha River (below US Route 25) Apalachee River Beaver Crk. (Taylor Co.) Brier Crk. (Burke Co.) Canoochee River (Hwy 192 to Lotts Crk.) Canoochee River (Lotts Crk. to Ogeechee River) Casey Canal Chattahoochee River (Helen to Lk. Lanier) (Buford Dam to Morgan Falls Dam) (Morgan Falls Dam to Peachtree Crk.) * (Peachtree Crk. to Pea Crk.) * (Pea Crk. to West Point Lk., below Franklin) * (West Point dam to I-85) (Oliver Dam to Upatoi Crk.) Chattooga River (NE Georgia, Rabun County) Chestatee River (below Tesnatee Riv.) Chickamauga Crk. (West) Cohulla Crk. (Whitfield Co.) Conasauga River (below Stateline) <18" Coosa River <20" 18 –32" (River Mile Zero to Hwy 100, Floyd Co.) ≥20" >32" <18" Coosa River <20" 18 –32" (Hwy 100 to Stateline, Floyd Co.) ≥20" >32" Coosa River (Coosa, Etowah below <20" Thompson-Weinman dam, Oostanaula) ≥20" Coosawattee River (below Carters) Etowah River (Dawson Co.) Etowah River (above Lake Allatoona) Etowah River (below Lake Allatoona dam) Flint River (Spalding/Fayette Cos.) Flint River (Meriwether/Upson/Pike Cos.) Flint River (Taylor Co.) Flint River (Macon/Dooly/Worth/Lee Cos.) <16" Flint River (Dougherty/Baker Mitchell Cos.) 16–30" >30" Gum Crk. -

Okefenokee Swamp Hydrology
OKEFENOKEE SWAMP HYDROLOGY Cynthia S. Loftin' AUTHOR: 'Graduate Research Assistant-Ph.D. Candidate, USGS-BRD Florida Cooperative Fish and Wildlife Research Unit, Department of Wildlife Ecology and Conservation, University of Florida, Gainesville, Florida 32611-0450. REFERENCE: Proceedings of the 1997 Georgia Water Resources Conference, held 20-22 March 1997, at the University of Georgia, Kathryn J. Hatcher, Editor, Institute of Ecology, The University of Georgia, Athens, Georgia. Abstract. The Okefenokee Swamp is one of North topographic relief is minimal The swamp is a bowl-like America's largest freshwater wetlands. Swamp hydrology depression in the landscape with the trend in ground surface is largely controlled by precipitation and elevation from 38.4 m at Kingfisher Landing in the evapotranspiration; regional topographic features of the Northeast to 33.0 m in the area where the Suwannee River swamp control surface water movements. Manipulations to exits the swamp in the West to 34.75 m at Ellicott's Mound the swamp topography and vegetation communities during in the Southeast near the St. Mary's River outflow. Within this century have affected water movement and variability the swamp are regional topographic highs on large sand- in parts of the swamp. Changes in swamp hydrology since based islands and lows in large prairies. The prairies also the construction of the Suwannee River Sill are generally contain local topographic highs on peat-based islands that restricted to the West Central area bounded by the Pocket, may rise a meter above the surrounding inundated peat Billy's Island, Craven's Island, Minnie's Island, and surface.