Characterization of Brain-Penetrant Pyrimidine-Containing Molecules
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Surnames and Migrations: the Barcelona Area (1451-1900)1
Surnames and Migrations: The Barcelona Area (1451-1900)1 Joan Pau Jordà Joana Maria Pujadas-Mora Anna Cabré Spain Abstract Catalan onomastics, and specifically the evolution of surnames, has been conditioned by several demographic, political and social processes that have imparted singular characteristics over the course of centuries. The combination of these factors resulted in a significant number of homonymic surnames, making it impossible to correctly identify their geographical origin based solely on linguistic criteria. As a possible solution to this, this paper proposes the use of the cluster analysis method to introduce a further criterion for their identification and classification. Historical registers of Marriage License Books from the Diocese of Barcelona are the source selected to achieve this goal. These records, which collect information on more than two million surnames, were maintained between 1451 and 1905 in a set of 291 books (Llibres d’Esposalles) kept at the archives of the Barcelona Cathedral. * * * Introduction The study of historical migrations is one of the most difficult demographic phenomena to investigate due to the absence of specific records until recent times. Given this lack, it is necessary to rely on indirect sources and methods that have already shown great potential, such as the analysis of surnames.2 However in the Catalan case – as well as in others – the evolution of onomastics, and specifically the evolution of surnames, has been conditioned by several demographic, political and social factors that have imparted singular characteristics over the course of centuries. The combination of these processes, as explained below, has made it necessary to propose the use of complementary methods to correctly identify the geographical origin of surnames and to complement existing linguistic criteria. -

Iain Crichton Smith 1928 - 1998
Iain Crichton Smith 1928 - 1998 Contents: Biography.................................................................................................................................................................Page 1 Two Old Women .....................................................................................................................................Pages 2 - 6 The End of An Auld Sang ....................................................................................................................Pages 6 - 8 The Beginning of a New Song....................................................................................................................Page 8 Further Reading / Contacts.............................................................................................................Pages 9 - 12 Biography: Iain Crichton Smith (1928 - 1998) (his gaelic name was Iain Mac a ‘Ghobhainn’) was born in Glasgow in 1928, Smith grew up from the age of two on the island of Lewis. The language of his upbringing was Gaelic; he learned English as his second language when he went to school at the Nicolson Institute in Stornoway. Later, he took a degree in English at the University of Aberdeen. From there he became a school teacher in Clydebank then Oban, where he could con- template his island upbringing at close range, but with a necessary degree of detachment. He retired early from teaching in 1977 to concentrate on his writing. Smith won various literary awards and was made an OBE in 1980. He published work in Gaelic under the -
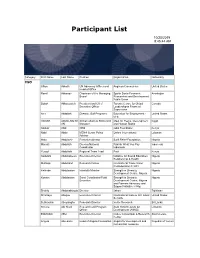
Participant List
Participant List 10/20/2019 8:45:44 AM Category First Name Last Name Position Organization Nationality CSO Jillian Abballe UN Advocacy Officer and Anglican Communion United States Head of Office Ramil Abbasov Chariman of the Managing Spektr Socio-Economic Azerbaijan Board Researches and Development Public Union Babak Abbaszadeh President and Chief Toronto Centre for Global Canada Executive Officer Leadership in Financial Supervision Amr Abdallah Director, Gulf Programs Educaiton for Employment - United States EFE HAGAR ABDELRAHM African affairs & SDGs Unit Maat for Peace, Development Egypt AN Manager and Human Rights Abukar Abdi CEO Juba Foundation Kenya Nabil Abdo MENA Senior Policy Oxfam International Lebanon Advisor Mala Abdulaziz Executive director Swift Relief Foundation Nigeria Maryati Abdullah Director/National Publish What You Pay Indonesia Coordinator Indonesia Yussuf Abdullahi Regional Team Lead Pact Kenya Abdulahi Abdulraheem Executive Director Initiative for Sound Education Nigeria Relationship & Health Muttaqa Abdulra'uf Research Fellow International Trade Union Nigeria Confederation (ITUC) Kehinde Abdulsalam Interfaith Minister Strength in Diversity Nigeria Development Centre, Nigeria Kassim Abdulsalam Zonal Coordinator/Field Strength in Diversity Nigeria Executive Development Centre, Nigeria and Farmers Advocacy and Support Initiative in Nig Shahlo Abdunabizoda Director Jahon Tajikistan Shontaye Abegaz Executive Director International Insitute for Human United States Security Subhashini Abeysinghe Research Director Verite -

Atomic Energy for Military Purposes (Princeton University Press, Princeton, New Jersey, 1945), Pp.98,143
REFERENCE 19 H. D. SMYTH, ATOMIC ENERGY FOR MILITARY PURPOSES (PRINCETON UNIVERSITY PRESS, PRINCETON, NEW JERSEY, 1945), PP.98,143. Atomic Energy for Military Purposes The Official Report on the Development of the Atomic Bomb under the Auspwes of the United States Government, 1940-l 945 By HENRY DEWOLF SMITH CHAIRMAN, DEPARTMENT OF PHYSICS PRINCETON UNIVERSITY CONSULTANT, MANHATT.4N DISTRICT, U.S. ENGINEERS Written at the request of MAJ. GEN. L. R. GROVES, U.S.-~. PRINCETON PRINCETON UNIVERSITY PRESS 1945 Copyright, 1945, by H. D. Smyth Reprociuctio~ in whoie 6 in p& authorized and permitted Printed in the United States of America , by Maple Press, York, Pennsylvania CHAPTER QI. T’E METALLURGICAL PROJECT AT CHICAGO IN 7942 INTRODUCTION 6.1, As has been made clear in Chapters IV and V, the infor- mation accumulated by the end of 1941 as to the possibility of producing an atomic bomb was such as to warrant expansion of the work, and this expansion called for an administrative reor- ganization. It was generally accepted that there was a very high probability that an atomic bomb of enormous destructive power could be made, either from concentrated U-235 or from the new element plutonium. It was proposed, therefore, to institute an intensive experimental and theoretical program including work both on isotope separation and on the chain-reaction problems. It was hoped that this program would establish definitely whether or not U-235 could be separated in significant quantities from U-238, either by electromagnetic or statistical methods; whether or not a chain reaction could be established with natural uranium or its compounds and could be made to yield relatively large quantities of plutonium; and whether or not the plutonium so produced could be separated from the parent material, uranium. -

North Carolina Association of Registers of Deeds
NORTH CAROLINA ASSOCIATION OF REGISTERS OF DEEDS MINIMUM STANDARDS FOR INDEXING REAL PROPERTY INSTRUMENTS Effective Date: July 1, 2012 (Except in Wake County in Which the Rules Shall Become Effective no Later Than 1/1/2014) 1 TABLE OF CONTENTS 1. STANDARD OF CARE AND POSTED INDEX RULES 1.01 Standard of Care in User Index Searches 1.02 Posting of Indexing Rules 2. CONSOLIDATED INDEX 2.01 Consolidated Real Property Index 3. INDEXING PROCEDURES 3.01 Spelling Variations and Illegibility 3.02 Additional Name References 3.03 Name Format 3.04 Names of All Parties 3.05 Basic Alphabetizing Procedures 3.06 Recognized Characters 3.07 Capital Letters 3.08 Names Required by General Statutes 4. NAMES 4.01 Surnames 4.02 Given Names 4.03 Titles 4.04 Suffixes 4.05 Surname in Parentheses 4.06 Professional Titles 4.07 Single Word Human Names 4.08 Estate Names 4.09 Trust Names 4.10 Non-Human Names 5. INDEX FORMAT 5.01 Sorting Order for Index 5.02 Trustee or Other Designated Status 5.03 Index Information 6. PLATS 6.01 Consolidated Real Property Index 6.02 Owner 6.03 Plat Title 6.04 Condominium Plat 7. MINIMUM FUNCTIONAL STANDARDS FOR ELECTONIC INDEXES 7.01 Compliance with Standards 7.02 Nonproprietary Data Storage and Accessibility 7.03 Sortable Fields 7.04 Search Result Narrowing Restrictions 7.05 Auto-Completion 7.06 Soundex Suggestions 7.07 Sorting Order 7.08 Allowable Name Length 7.09 Punctuation and Symbols 7.10 Hyphenated Surnames 7.11 Words Beginning The, A, An 7.12 Political Divisions 7.13 Abbreviations APPENDIX Recommended Instrument Type Abbreviations 2 1. -

Surname First Name Categorisation Abadin Jose Luis Silver Abbelen
2018 DRIVERS' CATEGORISATION LIST Updated on 09/07/2018 Drivers in red : revised categorisation Drivers in blue : new categorisation Surname First name Categorisation Abadin Jose Luis Silver Abbelen Klaus Bronze Abbott Hunter Silver Abbott James Silver Abe Kenji Bronze Abelli Julien Silver Abergel Gabriele Bronze Abkhazava Shota Bronze Abra Richard Silver Abreu Attila Gold Abril Vincent Gold Abt Christian Silver Abt Daniel Gold Accary Thomas Silver Acosta Hinojosa Julio Sebastian Silver Adam Jonathan Platinum Adams Rudi Bronze Adorf Dirk Silver Aeberhard Juerg Silver Afanasiev Sergei Silver Agostini Riccardo Gold Aguas Rui Gold Ahlin-Kottulinsky Mikaela Silver Ahrabian Darius Bronze Ajlani Karim Bronze Akata Emin Bronze Aksenov Stanislas Silver Al Faisal Abdulaziz Silver Al Harthy Ahmad Silver Al Masaood Humaid Bronze Al Qubaisi Khaled Bronze Al-Azhari Karim Bronze Alberico Neil Silver Albers Christijan Platinum Albert Michael Silver Albuquerque Filipe Platinum Alder Brian Silver Aleshin Mikhail Platinum Alesi Giuliano Silver Alessi Diego Silver Alexander Iradj Silver Alfaisal Saud Bronze Alguersuari Jaime Platinum Allegretta Vincent Silver Alleman Cyndie Silver Allemann Daniel Bronze Allen James Silver Allgàuer Egon Bronze Allison Austin Bronze Allmendinger AJ Gold Allos Manhal Bronze Almehairi Saeed Silver Almond Michael Silver Almudhaf Khaled Bronze Alon Robert Silver Alonso Fernando Platinum Altenburg Jeff Bronze Altevogt Peter Bronze Al-Thani Abdulrahman Silver Altoè Giacomo Silver Aluko Kolawole Bronze Alvarez Juan Cruz Silver Alzen -

Expressions of Existentialism and Absurdity in Gaelic Drama
The closed room: expressions of existentialism and absurdity in Gaelic drama Michelle Macleod, University of Aberdeen Introduction This paper is an exploration of six Gaelic plays written in the 1960s and 1970s: in particular it seeks to contextualise them with European drama of a roughly contemporaneous period by the likes of Ionesco, Sartre, Beckett and to demonstrate the internationalism of this genre of Gaelic writing. In the period under consideration Gaelic drama was most commonly performed as a one-act competition piece by amateur companies, (Macleod 2011) and the plays considered here were all part of that genre. While the Scottish Community Drama Association and An Comunn Gaidhealach competitions might seem far removed from Parisian and London theatres, the influence of the latter over the content of some of the former is visible. The articles in this journal and Macleod (2011) show that Gaelic drama has made a major contribution to the development of cultural expression within the Gaelic community on account of the volume of plays produced in amateur companies across the country and the number of people engaging actively with the arts. This paper, though not an exhaustive study of the Gaelic drama of this era, is a demonstration of the importance and excellence of this genre. It teases out various themes from Absurdist and existential theatre in other languages and uses some of the extensive scholarship of that drama as a mechanism to consider a selection of plays by three playwrights Finlay MacLeod (Fionnlagh MacLeòid) (1937-), Donnie Maclean (Donaidh MacIlleathain) (1939-2003) and Iain Crichton Smith (Iain Mac a’ Ghobhainn) (1928-1998), all from the Isle of Lewis. -

Case: 1:05-Cv-02746-SL Doc #: 132 Filed: 08/25/11 1 of 56. Pageid
Case: 1:05-cv-02746-SL Doc #: 132 Filed: 08/25/11 1 of 56. PageID #: <pageID> UNITED STATES DISTRICT COURT NORTHERN DISTRICT OF OHIO EASTERN DIVISION NANCY KOVACIC, ET AL., ) Case No. 1:05CV2746 ) Plaintiffs, ) JUDGE SARA LIOI ) v. ) ) MEMORANDUM OPINION CUYAHOGA COUNTY ) AND ORDER DEPARTMENT OF CHILDREN AND ) FAMILY SERVICES, et. al, ) ) Defendants. ) Plaintiffs in this case are Daniel and Katherine Kovacic, who were both minors when this suit was filed. Plaintiffs allege civil rights violations against various county and city entities and individual municipal employees and seek damages resulting from the warrantless entry into their home and their temporary removal from the care and custody of their mother, Nancy Kovacic. Following appeal and remand from the Sixth Circuit Court of Appeals, the remaining defendants include Cuyahoga County Department of Children and Family Services (CCDCFS), and social workers, Patricia Campbell Ponstingle, Pam Cameron, Vikki Csornok and Pam Gaylord (collectively, “defendants”). Before the Court are the motions for summary judgment filed by plaintiffs (Doc. 120) and defendants (Doc. 121). Defendants oppose plaintiffs’ motion (Doc. 125). Plaintiffs oppose defendants’ motion (Doc. 129), and defendants have filed a reply brief (Doc. 131). For the reasons that follow, the dispositive motions are GRANTED in part, and DENIED in part. I. BACKGROUND The history of this case dates back to 1995, when, following the contentious divorce of Nancy and Tom Kovacic, CCDCFS began receiving reports of alleged abuse and 1 Case: 1:05-cv-02746-SL Doc #: 132 Filed: 08/25/11 2 of 56. PageID #: <pageID> neglect by Nancy of the couple’s two children, Katherine and Daniel, including one incident where Daniel allegedly stabbed his mother with a pen. -

William E. Kovacic an Antitrust Tribute Liber Amicorum - Volume II
Editors Nicolas Charbit Elisa Ramundo Assistant Editors Anna M. Pavlik - Jessica Rebarber William E. Kovacic An Antitrust Tribute Liber Amicorum - Volume II Donald I. Baker, Jonathan B. Baker, Caron Beaton-Wells, Margaret Bloom, John DeQ. Briggs, George S. Cary, Andy C.M. Chen, Daniel A. Crane, Elaine Ewing, Eleanor M. Fox, Damien Geradin, Laurie-Anne Grelier, Omar Guerrero Rodríguez, Doris Hildebrand, Merit E. Janow, Joseph Kattan PC, Bruno Lasserre, Robert C. Marshall, Leslie M. Marx, Robert Ian McEwin, Andreas Mundt, Ali Nikpay, Maureen K. Ohlhausen, Julián Peña, Alan Ramírez Casazza, Patrick Rey, Simon Roberts, Jacques Steenbergen, John Terzaken, Thibaud Vergé, Florian Wagner-von Papp, Wouter P.J. Wils, Marc Winerman, Chris Wood, Joshua D. Wright. William E. Kovacic An Antitrust Tribute Liber Amicorum - Volume II Editors Nicolas Charbit Elisa Ramundo Assistant Editors Anna M. Pavlik - Jessica Rebarber © Institute of Competition Law, 2014 All rights reserved. No photocopying: copyright licenses do not apply. The information provided in this publication is general and may not apply in a specific situation. Legal advice should always be sought before taking any legal action based on the information provided. The publisher accepts no responsibility for any acts or omissions contained herein. Enquiries concerning reproduction should be sent to the Institute of Competition Law, at the address below. Copyright © 2014 by Institute of Competition Law 60 Broad Street, Suite 3502, NY 10004 www.concurrences.com [email protected] Printed in the United States of America First Printing, 2014 ISBN 978-1-939007-43-8 LCCN 2014940897 Publisher’s Cataloging-in-Publication (Provided by Quality Books, Inc.) William E. -

Names in English First Names and Surnames Personal Names
Names in English First names and surnames Personal names y Personal names in English are divided into first names, surnames and middle names. y An official name formula in England requires a first name,a surname,and a middle name, if there is one. y Legally, today it is possible to change a name in England and there is a special procedure for it. But, traditionally the change of one’s name wasn’t permitted without a permit of clerical authorities. y When they marry, women in England can choose between taking husband’s surname, leaving their maiden surname or adding husband’s surname to her surname. Names in Old English y When we speak about OE period we can talk about names only. There were no surnames. y Sources for the research of AS names were chronicles, inscriptions, legends, libri vitae, etc. One of the most important is Domesday book from 1086. It is a list of owners, users and values of lands in England after the Norman Conquest. y One of the main problems with texts from this period is the lack of a uniform ortography. Every scribe wrote in his own way. y Another very important work from earlier period is Historia Ecclesiastica Gentis Anglorum from 731, composed by a monk Venerable Bede. There are 215 names mentioned in it, but they are mostly aristocratic and the names of clergymen. AS names y An important fact for Anglosaxon names is that they had to be distinctive and each man could have only one name. This is why the naming system had to be able to produce a great number of different, fresh names. -

ACTA UNI VERSITATIS LODZIENSIS Piotr Stalmaszczyk “LANGUAGE IS
ACTA UNI VERSITATIS LODZIENSIS FOLIA LITTERARIA ANGLICA 7, 2007 Piotr Stalmaszczyk “LANGUAGE IS AT THE HEART OF MY WORK”: A NOTE ON THE POETRY OF IAIN CRICHTON SMITH (IAIN MAC A’GHOBHAINN) 1. INTRODUCTION One of the more interesting developments in twentieth-century Scottish Gaelic culture was the renaissance of poetry, especially in the second half of the century. Scottish Gaelic is spoken now by less than sixty thousand people, practically all of them English-speaking. However, in spite of this considerable decline in the number of Gaelic speakers, and to some extent also culture, the twentieth century saw a remarkable flowering of Gaelic literature, especially poetry.1 Traditional Gaelic poetry had an elaborate system of metres, it made use of end-rhyme, internal rhyme, alliteration, assonance, and introduced variation in line and stanza length. Early bardic verse in Ireland and Scotland observed a number of conventions and normative prescriptions, and bards were required to master specific linguistic knowledge necessary to construct appropriate verse. Modern Gaelic poetry differs from traditional in both form and content. Of the most important 20lh century Gaelic poets, George Campbell Hay (Deorsa Mac Iain Deorsa, 1915-1984) and Sorley MacLean (Somhairle MacGill-Eain, 1911-1996) worked within traditional metrical frameworks: George Campbell Hay revitalized traditional forms and created new elaborate sound-patterns, whereas Sorley MacLean crea tively transformed old patterns. Iain Crichton Smith (Iain Mac a’Ghobhainn, 1928-1998) often used regular length and rhyme but with variations of rhythm (a technique similar to the one used in his English poems). Derick ' Cf. the comments in the introductions to the collections of Gaelic poetry edited by MacAulay (1976), Davitt and MacDhomhnaill (1993), and Black (1999). -

Cayuga County Surname and Family Files in Town and Organization Collections
Cayuga County Surname and Family Files In town and organization collections A B C D E F G 1 Surname Cayuga County Genoa Moravia Town of Town of Montezuma 2 x = family name on file Historian Hist. Assn. COLHS Sterling Victory Hist. Society 3 4 Moravia names also online @ www.colhs.org/p/surnames see 5 Montezuma names online @ www.montezumagen.com website 6 7 Abbe x 8 Abbott x x x x 9 Abbott-Nuitt x 10 Abraham(s) x 11 Abrams x x x 12 Acers x 13 Acker x 14 Ackerman x x 15 Ackerson x x x 16 Ackles x 17 Ackley x 18 Acre x 19 Adams x x x x 20 Adams-Crofoot x 21 Addy x 22 Adessa x 23 Adkins x 24 Adle x 25 Adolph x 26 Adriance x x 27 Adsitt x 28 Agree x 29 Aiken/Aikin x 30 Aikin x 31 Akin x 32 Akin (Aiken) x 33 Albertson x 34 Albie/Albee x 35 Albring x 36 Albro x 37 Alcorn x 38 Alcott/Alcox x 39 Alden x 40 Aldrich x x 41 Aldridge x 42 Alexander x x 43 Alfred x 44 Alger/Algur x 45 Algert x 46 Alifieri x 47 Alissandrello x 48 Allanson x 49 Allee x 50 Allen x x x x x 51 Alley x 52 Allis x 53 Almy x 54 Alnutt x x Cayuga County Surname and Family Files In town and organization collections A B C D E F G 1 Surname Cayuga County Genoa Moravia Town of Town of Montezuma 2 x = family name on file Historian Hist.