Cyathus Lignilantanae Sp. Nov., a New Species of Bird's Nest Fungi
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Why Mushrooms Have Evolved to Be So Promiscuous: Insights from Evolutionary and Ecological Patterns
fungal biology reviews 29 (2015) 167e178 journal homepage: www.elsevier.com/locate/fbr Review Why mushrooms have evolved to be so promiscuous: Insights from evolutionary and ecological patterns Timothy Y. JAMES* Department of Ecology and Evolutionary Biology, University of Michigan, Ann Arbor, MI 48109, USA article info abstract Article history: Agaricomycetes, the mushrooms, are considered to have a promiscuous mating system, Received 27 May 2015 because most populations have a large number of mating types. This diversity of mating Received in revised form types ensures a high outcrossing efficiency, the probability of encountering a compatible 17 October 2015 mate when mating at random, because nearly every homokaryotic genotype is compatible Accepted 23 October 2015 with every other. Here I summarize the data from mating type surveys and genetic analysis of mating type loci and ask what evolutionary and ecological factors have promoted pro- Keywords: miscuity. Outcrossing efficiency is equally high in both bipolar and tetrapolar species Genomic conflict with a median value of 0.967 in Agaricomycetes. The sessile nature of the homokaryotic Homeodomain mycelium coupled with frequent long distance dispersal could account for selection favor- Outbreeding potential ing a high outcrossing efficiency as opportunities for choosing mates may be minimal. Pheromone receptor Consistent with a role of mating type in mediating cytoplasmic-nuclear genomic conflict, Agaricomycetes have evolved away from a haploid yeast phase towards hyphal fusions that display reciprocal nuclear migration after mating rather than cytoplasmic fusion. Importantly, the evolution of this mating behavior is precisely timed with the onset of diversification of mating type alleles at the pheromone/receptor mating type loci that are known to control reciprocal nuclear migration during mating. -

Tree of Life Marula Oil in Africa
HerbalGram 79 • August – October 2008 HerbalGram 79 • August Herbs and Thyroid Disease • Rosehips for Osteoarthritis • Pelargonium for Bronchitis • Herbs of the Painted Desert The Journal of the American Botanical Council Number 79 | August – October 2008 Herbs and Thyroid Disease • Rosehips for Osteoarthritis • Pelargonium for Bronchitis • Herbs of the Painted Desert • Herbs of the Painted Bronchitis for Osteoarthritis Disease • Rosehips for • Pelargonium Thyroid Herbs and www.herbalgram.org www.herbalgram.org US/CAN $6.95 Tree of Life Marula Oil in Africa www.herbalgram.org Herb Pharm’s Botanical Education Garden PRESERVING THE FULL-SPECTRUM OF NATURE'S CHEMISTRY The Art & Science of Herbal Extraction At Herb Pharm we continue to revere and follow the centuries-old, time- proven wisdom of traditional herbal medicine, but we integrate that wisdom with the herbal sciences and technology of the 21st Century. We produce our herbal extracts in our new, FDA-audited, GMP- compliant herb processing facility which is located just two miles from our certified-organic herb farm. This assures prompt delivery of freshly-harvested herbs directly from the fields, or recently HPLC chromatograph showing dried herbs directly from the farm’s drying loft. Here we also biochemical consistency of 6 receive other organic and wildcrafted herbs from various parts of batches of St. John’s Wort extracts the USA and world. In producing our herbal extracts we use precision scientific instru- ments to analyze each herb’s many chemical compounds. However, You’ll find Herb Pharm we do not focus entirely on the herb’s so-called “active compound(s)” at fine natural products and, instead, treat each herb and its chemical compounds as an integrated whole. -

Cuivre Bryophytes
Trip Report for: Cuivre River State Park Species Count: 335 Date: Multiple Visits Lincoln County Agency: MODNR Location: Lincoln Hills - Bryophytes Participants: Bryophytes from Natural Resource Inventory Database Bryophyte List from NRIDS and Bruce Schuette Species Name (Synonym) Common Name Family COFC COFW Acarospora unknown Identified only to Genus Acarosporaceae Lichen Acrocordia megalospora a lichen Monoblastiaceae Lichen Amandinea dakotensis a button lichen (crustose) Physiaceae Lichen Amandinea polyspora a button lichen (crustose) Physiaceae Lichen Amandinea punctata a lichen Physiaceae Lichen Amanita citrina Citron Amanita Amanitaceae Fungi Amanita fulva Tawny Gresette Amanitaceae Fungi Amanita vaginata Grisette Amanitaceae Fungi Amblystegium varium common willow moss Amblystegiaceae Moss Anisomeridium biforme a lichen Monoblastiaceae Lichen Anisomeridium polypori a crustose lichen Monoblastiaceae Lichen Anomodon attenuatus common tree apron moss Anomodontaceae Moss Anomodon minor tree apron moss Anomodontaceae Moss Anomodon rostratus velvet tree apron moss Anomodontaceae Moss Armillaria tabescens Ringless Honey Mushroom Tricholomataceae Fungi Arthonia caesia a lichen Arthoniaceae Lichen Arthonia punctiformis a lichen Arthoniaceae Lichen Arthonia rubella a lichen Arthoniaceae Lichen Arthothelium spectabile a lichen Uncertain Lichen Arthothelium taediosum a lichen Uncertain Lichen Aspicilia caesiocinerea a lichen Hymeneliaceae Lichen Aspicilia cinerea a lichen Hymeneliaceae Lichen Aspicilia contorta a lichen Hymeneliaceae Lichen -

Fungal Planet Description Sheets: 716–784 By: P.W
Fungal Planet description sheets: 716–784 By: P.W. Crous, M.J. Wingfield, T.I. Burgess, G.E.St.J. Hardy, J. Gené, J. Guarro, I.G. Baseia, D. García, L.F.P. Gusmão, C.M. Souza-Motta, R. Thangavel, S. Adamčík, A. Barili, C.W. Barnes, J.D.P. Bezerra, J.J. Bordallo, J.F. Cano-Lira, R.J.V. de Oliveira, E. Ercole, V. Hubka, I. Iturrieta-González, A. Kubátová, M.P. Martín, P.-A. Moreau, A. Morte, M.E. Ordoñez, A. Rodríguez, A.M. Stchigel, A. Vizzini, J. Abdollahzadeh, V.P. Abreu, K. Adamčíková, G.M.R. Albuquerque, A.V. Alexandrova, E. Álvarez Duarte, C. Armstrong-Cho, S. Banniza, R.N. Barbosa, J.-M. Bellanger, J.L. Bezerra, T.S. Cabral, M. Caboň, E. Caicedo, T. Cantillo, A.J. Carnegie, L.T. Carmo, R.F. Castañeda-Ruiz, C.R. Clement, A. Čmoková, L.B. Conceição, R.H.S.F. Cruz, U. Damm, B.D.B. da Silva, G.A. da Silva, R.M.F. da Silva, A.L.C.M. de A. Santiago, L.F. de Oliveira, C.A.F. de Souza, F. Déniel, B. Dima, G. Dong, J. Edwards, C.R. Félix, J. Fournier, T.B. Gibertoni, K. Hosaka, T. Iturriaga, M. Jadan, J.-L. Jany, Ž. Jurjević, M. Kolařík, I. Kušan, M.F. Landell, T.R. Leite Cordeiro, D.X. Lima, M. Loizides, S. Luo, A.R. Machado, H. Madrid, O.M.C. Magalhães, P. Marinho, N. Matočec, A. Mešić, A.N. Miller, O.V. Morozova, R.P. Neves, K. Nonaka, A. Nováková, N.H. -

Redalyc.Novelties of Gasteroid Fungi, Earthstars and Puffballs, from The
Anales del Jardín Botánico de Madrid ISSN: 0211-1322 [email protected] Consejo Superior de Investigaciones Científicas España da Silva Alfredo, Dönis; de Oliveira Sousa, Julieth; Jacinto de Souza, Elielson; Nunes Conrado, Luana Mayra; Goulart Baseia, Iuri Novelties of gasteroid fungi, earthstars and puffballs, from the Brazilian Atlantic rainforest Anales del Jardín Botánico de Madrid, vol. 73, núm. 2, 2016, pp. 1-10 Consejo Superior de Investigaciones Científicas Madrid, España Available in: http://www.redalyc.org/articulo.oa?id=55649047009 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative Anales del Jardín Botánico de Madrid 73(2): e045 2016. ISSN: 0211-1322. doi: http://dx.doi.org/10.3989/ajbm.2422 Novelties of gasteroid fungi, earthstars and puffballs, from the Brazilian Atlantic rainforest Dönis da Silva Alfredo1*, Julieth de Oliveira Sousa1, Elielson Jacinto de Souza2, Luana Mayra Nunes Conrado2 & Iuri Goulart Baseia3 1Programa de Pós-Graduação em Sistemática e Evolução, Centro de Biociências, Campus Universitário, 59072-970, Natal, RN, Brazil; [email protected] 2Curso de Graduação em Ciências Biológicas, Universidade Federal do Rio Grande do Norte, Campus Universitário, 59072-970, Natal, Rio Grande do Norte, Brazil 3Departamento de Botânica e Zoologia, Universidade Federal do Rio Grande do Norte, Campus Universitário, 59072970, Natal, Rio Grande do Norte, Brazil Recibido: 24-VI-2015; Aceptado: 13-V-2016; Publicado on line: 23-XII-2016 Abstract Resumen Alfredo, D.S., Sousa, J.O., Souza, E.J., Conrado, L.M.N. -

Revision of the Genus Cyathus (Basidiomycota) from the Herbaria of Northeast Brazil
Mycosphere 5 (4): 531–540 (2014) ISSN 2077 7019 www.mycosphere.org Article Mycosphere Copyright © 2014 Online Edition Doi 10.5943/mycosphere/5/4/5 Revision of the genus Cyathus (Basidiomycota) from the herbaria of northeast Brazil Cruz RHSF1, Assis NM2, Silva MA3 and Baseia IG4 1Programa de Pós-Graduação em Sistemática e Evolução, Centro de Biociências, Universidade Federal do Rio Grande do Norte, Avenida Senador Salgado Filho, 3000, Natal-RN 59.078-970 Brazil, [email protected] 2Departamento de Botânica e Zoologia, Centro de Biociências, Universidade Federal do Rio Grande do Norte, Avenida Senador Salgado Filho, 3000, Natal-RN 59.078-970 Brazil, [email protected] 3Departamento de Micologia, Universidade Federal de Pernambuco, Avenida Professor Moraes Rego 1235, Recife-PE 50.670-901 Brazil, [email protected] 4Departamento de Botânica e Zoologia, Centro de Biociências, Universidade Federal do Rio Grande do Norte, Avenida Senador Salgado Filho, 3000, Natal-RN 59.078-970 Brazil, [email protected] Cruz RHSF, Assis NM, Silva MA, Baseia IG 2014 – Revision of the genus Cyathus (Basidiomycota) from the herbaria of northeast Brazil. Mycosphere 5(4), 531−540, Doi 10.5943/mycosphere/5/4/5 Abstract Seventy exsiccates of the genus Cyathus deposited in JPB, UESC, URM and UFRN herbaria were studied and nine species were identified: Cyathus badius, C. berkeleyanus, C. earlei, C. gracilis, C. limbatus, C. pallidus, C. poeppigii, C. setosus and C. striatus. Cyathus berkeleyanus and C. poeppigii are recorded for the first time for northeastern Brazil. Descriptions, taxonomic remarks and illustrations of the studied material are presented. Key words – herbarium collection – Nidulariaceae – Gasteromycetes – taxonomic review Introduction The genus Cyathus Haller belongs to the family Nidulariaceae, included in the agaricoid clade of Basidiomycota (Matheny et al. -

Identification and Management of Heart-Rot Fungi
https://doi.org/10.3126/banko.v30i2.33482 Banko Janakari, Vol 30 No. 2, 2020 Pp 71‒77 Short Note Identification and management of heart-rot fungi S. K. Jha1 eart-rot fungi are key players in trees moist, soft, spongy, or stringy and appear white health, diversity and nutrient dynamic or yellow. Mycelia of fungi colonize much of Hin forest as pathogens and decomposers the woody tissues. White rots usually form in along with a number of invertebrates are flowering trees (angiosperms) and less often in associated with Wood-decay fungi serve as conifers (gymnosperms). Fungi that cause white vectors for fungal pathogens, or are fungivorous rots also cause the production of zone lines in and influence rates of Wood-decay and nutrient wood, sometimes called "spalted wood". This mineralization. partially rotted wood is sometimes desirable for woodworking. The examples of white rot fungi A number of fungi, viz. Polyporus spp., Serpulala are Armillariell amellea, Pleurotus ostreatus, crymans, Fusarium negundi, Coniophora Coriolus versicolor, Cyathus stercoreus, cerebella, Lentinus lapidens and Penicillium Ceriporiopsissu bvermispora, Trametes divaricatum cause destruction of valuable versicolor, Hetero basidionannosum, and so on. timbers by reducing the mechanical strength of wood. Molds cause rotting of the heartwood in Brown rots the middle of tree-branches and trunks. Wood- decay fungi can be classified according to the Brown rots primarily decay the cellulose and type of decay that they cause. The best-known hemicellulose (carbohydrates) in wood, leaving types are brown rot, soft rot, and white rot. Each behind the brownish lignin. Wood affected by type produces different enzymes, can degrade brown rot usually is dry, fragile, and readily different plant materials, and can colonize crumbles into cubes because of longitudinal different environmental niches (Bednarz et al. -

Gasteroid Mycobiota (Agaricales, Geastrales, And
Gasteroid mycobiota ( Agaricales , Geastrales , and Phallales ) from Espinal forests in Argentina 1,* 2 MARÍA L. HERNÁNDEZ CAFFOT , XIMENA A. BROIERO , MARÍA E. 2 2 3 FERNÁNDEZ , LEDA SILVERA RUIZ , ESTEBAN M. CRESPO , EDUARDO R. 1 NOUHRA 1 Instituto Multidisciplinario de Biología Vegetal, CONICET–Universidad Nacional de Córdoba, CC 495, CP 5000, Córdoba, Argentina. 2 Facultad de Ciencias Exactas Físicas y Naturales, Universidad Nacional de Córdoba, CP 5000, Córdoba, Argentina. 3 Cátedra de Diversidad Vegetal I, Facultad de Química, Bioquímica y Farmacia., Universidad Nacional de San Luis, CP 5700 San Luis, Argentina. CORRESPONDENCE TO : [email protected] *CURRENT ADDRESS : Centro de Investigaciones y Transferencia de Jujuy (CIT-JUJUY), CONICET- Universidad Nacional de Jujuy, CP 4600, San Salvador de Jujuy, Jujuy, Argentina. ABSTRACT — Sampling and analysis of gasteroid agaricomycete species ( Phallomycetidae and Agaricomycetidae ) associated with relicts of native Espinal forests in the southeast region of Córdoba, Argentina, have identified twenty-nine species in fourteen genera: Bovista (4), Calvatia (2), Cyathus (1), Disciseda (4), Geastrum (7), Itajahya (1), Lycoperdon (2), Lysurus (2), Morganella (1), Mycenastrum (1), Myriostoma (1), Sphaerobolus (1), Tulostoma (1), and Vascellum (1). The gasteroid species from the sampled Espinal forests showed an overall similarity with those recorded from neighboring phytogeographic regions; however, a new species of Lysurus was found and is briefly described, and Bovista coprophila is a new record for Argentina. KEY WORDS — Agaricomycetidae , fungal distribution, native woodlands, Phallomycetidae . Introduction The Espinal Phytogeographic Province is a transitional ecosystem between the Pampeana, the Chaqueña, and the Monte Phytogeographic Provinces in Argentina (Cabrera 1971). The Espinal forests, mainly dominated by Prosopis L. -

Fluted Bird's Nest Fungus, Cyathus Striatus
A Horticulture Information article from the Wisconsin Master Gardener website, posted 19 Sept 2014 Fluted Bird’s Nest Fungus, Cyathus striatus There are many fungi in several genera called bird’s nest fungi because of the resemblance of their fruiting bodies to a tiny nest fi lled with eggs. One of the most common in Wisconsin is Cyathus striatus, the fl uted bird’s nest fungus. This species is widespread throughout temperate regions of the world, developing on dead wood in open forests, typically growing individually or in clusters on small twigs and fallen branches or other wood debris. Because it also grows readily in bark or wood mulch, it is frequently found in landscaped yards and gardens. Other species grow on plant remains or cow or horse dung. C. striatus, and others, are most commonly Fruiting bodies of fl uted bird’s nest seen in the autumn fungus, Cyathus striatus. when damp conditions promote their development, but they can be seen anytime conditions are appropriate. Even though each individual is small and inconspicuous, this species often grows in huge clusters, making A large cluster of fl uted bird’s nest fungi growing them more noticeable – on bark mulch. although they blend in so well with their background that it is very easy to overlook them. All of the bird’s nest fungi look like miniature nests (generally only ¼ inch in diameter) fi lled with four or fi ve tiny eggs. The cup-shaped “nest”, called a peridium, may be brown, gray or white, and smooth or textured inside and out. -

A New Species of Bird's Nest Fungi: Characterisation of <I>Cyathus Subglobisporus</I>
Persoonia 21, 2008: 71–76 www.persoonia.org RESEARCH ARTICLE doi:10.3767/003158508X370578 A new species of bird’s nest fungi: characterisation of Cyathus subglobisporus sp. nov. based on morphological and molecular data R.-L. Zhao1, D.E. Desjardin 2, K. Soytong 3, K.D. Hyde 4, 5* Key words Abstract Recent collections of bird’s nest fungi (i.e. Crucibulum, Cyathus, Mycocalia, Nidula, and Nidularia species) in northern Thailand resulted in the discovery of a new species of Cyathus, herein described as C. subglobisporus. bird’s nest fungi This species is distinct by a combination of ivory-coloured fruiting bodies covered with shaggy hairs, plications on gasteromycetes the inner surface of the peridium and subglobose basidiospores. Phylogenetic analyses based on ITS and LSU new species ribosomal DNA sequences using neighbour-joining, maximum likelihood and weighted maximum parsimony sup- phylogeny port Cyathus subglobisporus as a distinct species and sister to a clade containing C. annulatus, C. renweii and rDNA C. stercoreus in the Striatum group. Article info Received: 24 June 2008; Accepted: 8 September 2008; Published: 23 September 2008. INTRODUCTION Cyathus striatus as representatives of the Nidulariaceae. Their phylogenetic reconstruction indicated that the Nidulariaceae The genus Cyathus along with the genera Crucibulum, Myco- was sister to the Cystodermateae (represented by Cystoderma calia, Nidula, and Nidularia are known as the bird’s nest fungi amianthinum). Together these two clades appear sister to the because of their small vase-shaped or nest-like fruiting bodies Agaricaceae s.l. but without bootstrap support. A phylogenetic containing lentil-shaped or egg-like peridioles. -

Allescheria Boydii Shear
OBSERVATIONS ON THE MORPHOLCGY, PHYSIOLCGY, AND LIFE HISTORY OF ALLESCHERIA BOYDII SHEAR. by Robert Denton Flory A Thesis Presented to the Graduate School of the University of Richmond in Partial Fulfillment of the Requirements for the Degree of Master of Arts University of Richmond June, 1956 LlBRARY UNIVERSITY OF RICHMOND VIRGINIA ACKNOWLEDGEMENTS To those individuals who have contributed to the success of this thesis, I wish to express rrry sincere appreciation. I am especially grateful to Dr. R. F. Smart, who made possible for me the opportunity to study at the University of Richmond, and for his constant guidance throughout the preparation of this thesis. I am also greatly indebted to the other members of the faculty at the University of Richmond Biology Department for their advice and for the many considerations they have shown. It is a pleasure to acknowledge the aid of Dr. C. W. Emmons of the National Institute of Health for making available many strains of the fungus studied. Grateful acknowledgement is made to Dr. R. F. Smart, Dr •. J. D. Burke, Mr. A.H. O'Bier, Jr. and Mr. R. F. Moore,Jr,. for their assistance with the photographic plates of this thesis. LIBRARY °lJNlVERSlTY OF RICHMOND VIRGINIA II CONTENTS page ACl<l'iOWI.EDG~S. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • II IN'rRODUCTI ON •• ·• • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 1 ~I.AI.iS .AND J.i:E'rHODS. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 3 RESULTS OF MORPHOLOGIC.AL .AND CYTOLOGICAL STUDY ••••••••••• 6 Colony Characteristics ••••••••••••••.••••••••••••••• 6 Vegetative Hyphae •••.•.•••••.••••.••.•••.••••••••••. 8 Asexual Reprod.uction. • . • . • • . • • • • • . • • • • • . • • • • • • • • . • • l2 Sexual Reproduction ••••.•••••••••••••••• 18 PHYSIC.AL EFFECTS OF ENVIRONMENT UPON GROWTH. • . • • • • • • • • • • • 28 NUTRITIONAL STUDIES .AND OCCURRENCE IN NATURE •••••..•••.•• 32 LIFE HISTORY, SEXUALITY, .AND TAXONOMY Life History •• . -
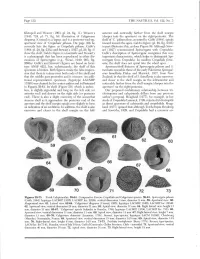
(1943: 724, Pi. 71, Fig. 16) Illustration of Calyptraea (Deeper Into the Aperture) on the Right/Posterior
Page 118 THE NAUTILUS, Vol. 122, No. 3 Kleinpeil and Weaver (1963, pi. 24, fig. 11). Weavers anterior and noticeably farther from the shell margin (1943: 724, pi. 71, fig. 16) illustration of Calyptraea (deeper into the aperture) on the right/posterior. The diegoana (Conrad) is a lapsus and is a posterior-end-up, shelf of 'C.y pileum thus, as noted by Gabb (1864), spirals apertural view of 'Crepidula' pileum. On page 356 he inward toward the apex. Gabb's figure (pi. 29, fig. 233b) correctly lists the figure as Crepidula pileum. Gabb's in part illustrates this, as does Figure 59. Although Stew- (1864: pi. 29, fig. 233a) and Stewart's (1927, pi. 29, fig. 3) art (1927) synonomized Spirocrypta with Crepidula, show the shelf. Gabb's figure is a fascimile and Stewart's Gabb's description of Spirocrypta recognizes this very is a photograph that has been reproduced in other dis- important characteristic, which helps to distinguish Spi- cussions of Spirocrypta (e.g., Wenz, 1940: 903, fig. rocrypta from Crepidula. In modern Crepidula forni- 2660a). Gabb's and Stewart's figures are based on lecto- cata, the shelf does not spiral into the whorl apex. type ANSP 4221, but, unfortunately, the shelf of this Aperture/shelf features of Spirocrypta pileum and S. specimen is broken. Both figures create the false impres- inomata resemble those of the early Paleocene Spirogal- sion that there is a sinus near both ends of the shelf and erus lamellaria Finlay and Marwick, 1937, from New that the middle part protrudes and is concave.