OCCURRENCE of SIEVE ELEMENTS in PHLOEM RAYS Kishore S. Rajput & K. S. Rao Solitary Sieve Elements Or Groups of Sieve Element
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bark Anatomy and Cell Size Variation in Quercus Faginea
Turkish Journal of Botany Turk J Bot (2013) 37: 561-570 http://journals.tubitak.gov.tr/botany/ © TÜBİTAK Research Article doi:10.3906/bot-1201-54 Bark anatomy and cell size variation in Quercus faginea 1,2, 2 2 2 Teresa QUILHÓ *, Vicelina SOUSA , Fatima TAVARES , Helena PEREIRA 1 Centre of Forests and Forest Products, Tropical Research Institute, Tapada da Ajuda, 1347-017 Lisbon, Portugal 2 Centre of Forestry Research, School of Agronomy, Technical University of Lisbon, Tapada da Ajuda, 1349-017 Lisbon, Portugal Received: 30.01.2012 Accepted: 27.09.2012 Published Online: 15.05.2013 Printed: 30.05.2013 Abstract: The bark structure of Quercus faginea Lam. in trees 30–60 years old grown in Portugal is described. The rhytidome consists of 3–5 sequential periderms alternating with secondary phloem. The phellem is composed of 2–5 layers of cells with thin suberised walls and narrow (1–3 seriate) tangential band of lignified thick-walled cells. The phelloderm is thin (2–3 seriate). Secondary phloem is formed by a few tangential bands of fibres alternating with bands of sieve elements and axial parenchyma. Formation of conspicuous sclereids and the dilatation growth (proliferation and enlargement of parenchyma cells) affect the bark structure. Fused phloem rays give rise to broad rays. Crystals and druses were mostly seen in dilated axial parenchyma cells. Bark thickness, sieve tube element length, and secondary phloem fibre wall thickness decreased with tree height. The sieve tube width did not follow any regular trend. In general, the fibre length had a small increase toward breast height, followed by a decrease towards the top. -

Parenchyma Cell Respiration and Survival in Secondary Xylem: Does Metabolic Activity Decline with Cell Age?
Plant, Cell and Environment (2007) 30, 934–943 doi: 10.1111/j.1365-3040.2007.01677.x Parenchyma cell respiration and survival in secondary xylem: does metabolic activity decline with cell age? R. SPICER1 & N. M. HOLBROOK2 1Rowland Institute at Harvard University, 100 Edwin H. Land Boulevard, Cambridge, MA 02142, USA and 2Organismic and Evolutionary Biology, Harvard University, 16 Divinity Avenue, Cambridge, MA 02138, USA ABSTRACT defines (and arguably drives) heartwood formation, a form of tissue senescence during which the oldest, non- Sapwood respiration often declines towards the sapwood/ functional xylem is compartmentalized in the centre of the heartwood boundary, but it is not known if parenchyma stem. The cause of parenchyma cell death is not known, metabolic activity declines with cell age. We measured but evidence for decreased metabolic activity in the inner- sapwood respiration in five temperate species (sapwood age most sapwood has led to a view of parenchyma ageing as range of 5–64 years) and expressed respiration on a live cell a gradual, passive decline in metabolism that terminates in basis by quantifying living parenchyma. We found no effect cell death. of parenchyma age on respiration in two conifers (Pinus Multiple reports suggest that sapwood respiration strobus, Tsuga canadensis), both of which had signifi- declines towards the sapwood/heartwood boundary cant amounts of dead parenchyma in the sapwood. In (Goodwin & Goddard 1940; Higuchi, Shimada & Watanabe angiosperms (Acer rubrum, Fraxinus americana, Quercus 1967; Pruyn, Gartner & Harmon 2002a,b; Pruyn, Harmon & rubra), both bulk tissue and live cell respiration were Gartner 2003; Pruyn, Gartner & Harmon 2005), although reduced by about one-half in the oldest relative to the there have been reports of no change (Bowman et al. -

Plant Histology and Anatomy Q
PLANT HISTOLOGY AND ANATOMY Q. Transcellular strands are seen in a) Xylem vessels b) ThidTracheids c) Parenchyma cells d) Sieve tubes Q. Epiphytes absorb water by a spongy tissue called a) Mesophy ll b) Velamen c) Conjuctive tissue d) Phloem Q. The presence of vessel s and companion cells are characters of a) Gymnosperms b) Angiosperms c) Bryophytes d) Pteridophytes Q. AhilAmphivasal vascul lbdlar bundle is found in a) Cycas and Dryopteris b) Dracaena and Yucca c) Helianthus and Cucurbita d) maize and Wheat Q. Bamboo and grasses elonggyate by the activity of a) Apical meristem b) Intercalary meristem c) SdSecondary meritistem d) Lateral ameristem Q. Fibres associated with phloem are called a) Intraxylary fibres b) Pericycle fibres c) Bast fibres d) Cortical fibres Q. Callose is found in a) Sieve Plates b) Cross walls of tracheids c) Phloem parenchyma d) Comapanion cell Q. Which are common in xylem and phloem tissues? a) PhParenchyma and CllCollench yma b) Collenchyma and Sclerenchyma c) Parenchyma and Sclerenchyma d) Aerenchyma and Sclerenchyma Q. Quiescent centre is found in a) Root tip b) Shoot tip c) Floral tip d) Leaf tip Q. The plastids in meristematic tissue are in a a) Fullyyp developed state b) Half developed state c) Proplastid state d) Plasmolysed state Q. In hydrophytes, aerenchyma helps in a) Attachment b) Photosynthesis c) Buoyancy d) Mechanical support Q. Cistoliths are composedfd of a) Calcium oxalate b) Calcium carbonate c) GGucosdeslucosides d) MgCO 3 Q. CllCollench yma differs f rom sclerenchyma a) Retaining protoplasm at maturity b) Having thick walls c) HHiaving w idlide lumen d) Being meristematic Q. -

Microbiology and Plant Pathology
ALAGAPPA UNIVERSITY (Accredited with ‘A+’ Grade by NAAC (with CGPA: 3.64) in the Third Cycle and Graded As category - I University by MHRD-UGC) (A State University Established by the Government of TamilNadu) KARAIKUDI – 630 003 DIRECTORATE OF DISTANCE EDUCATION M. Sc. BOTONY Second Year – Third Semester 34631- MICROBIOLOGY AND PLANT PATHOLOGY Copy Right Reserved For Private Use only Authors: Dr. A. Arun, Associate Professor & Head (i/c), Department of Microbiology, Alagappa University, Karaikudi- 630 003. (Units 1-7) Dr. M. Jothibasu Assistant Professor Department of Botany Alagappa University Karaikudi-03 (Units 8-14) “The Copyright shall be vested with Alagappa University” All rights reserved. No part of this publication which is material protected by this copyright notice may be reproduced or transmitted or utilized or stored in any form or by any means now known or hereinafter invented, electronic, digital or mechanical, including photocopying, scanning, recording or by any information storage or retrieval system, without prior written permission from the Alagappa University, Karaikudi, Tamil Nadu. SYLLABI – BOOK MAPPING TABLE 34631- MICROBIOLOGY AND PLANT PATHOLOGY SYLLABI MAPPING IN BOOK BLOCK-1: SCOPE OF MICROBIOLOGY UNIT I Introduction to Microbiology- Scope of Microbiology- 1-8 Evolution into Science- Characterization of microorganisms. UNIT II Classification of Microorganisms- Archea and Bacteria- 9-15 Characteristics of Bacteria- Morphology -Ultra Structure. UNIT III Nutrition – Growth – Reproduction- Bacterial Cultural and 16-27 Cultural characteristics- Economic importance of bacteria BLOCK 2: VIRUS AND MYCOPLASMAS UNIT IV Virology – General features- Classification of virus- 28-34 Characteristics – Ultra structure. UNIT V Virus- Isolation – Purification –Chemical nature- Replication- 35-46 Transmission- Virions. -

Does the Distance to Normal Renal Parenchyma (DTNRP) in Nephron-Sparing Surgery for Renal Cell Carcinoma Have an Effect on Survival?
ANTICANCER RESEARCH 25: 1629-1632 (2005) Does the Distance to Normal Renal Parenchyma (DTNRP) in Nephron-sparing Surgery for Renal Cell Carcinoma have an Effect on Survival? Z. AKÇETIN1, V. ZUGOR1, D. ELSÄSSER1, F.S. KRAUSE1, B. LAUSEN2, K.M. SCHROTT1 and D.G. ENGEHAUSEN1 Departments of 1Urology and 2Medical Informatics, Biometry and Epidemiology, University of Erlangen-Nuremberg, Germany Abstract. Background: The effect of the distance to normal renal solitary kidneys. Additionally, organ preservation in the parenchyma (DTNRP) on survival after nephron-sparing surgery presence of an intact contralateral kidney can be performed (NSS) for renal cell cancer (RCC) was analyzed. Additionally, for small localized tumors with nearly equivalent results for the role of T-classification, tumor diameter and tumor grading tumor-specific survival, compared to nephrectomy (1). The was considered. Patients and Methods: NSS was performed on question of whether a small safety margin in intraoperative 126 patients with RCC between 1988 and 2000. Eighty-six patients histology may be adequate for favorable outcome of the were submitted to annual follow-up. These 86 patients were sub- patient constitutes an everyday issue for the practitioner classified into statistical groups according to the distance performing nephron-sparing surgery. In this context, the to normal renal parenchyma (≤ 2mm; > 2mm – ≤ 5mm; clinical impact of defined surgical margin widths for >5 mm), T-classification, tumor diameter (≤ 20mm; > 20mm - avoiding local tumor recurrence and, therefore, improved ≤ 30 mm; >30 mm – ≤ 50mm; >50mm) and tumor grading. survival after nephron-sparing surgery has been discussed The effect of belonging to one of these groups on survival was but still remains controversial. -

INTERXYLARY PHLOEM (Included Phloem) by Marcelo R
INTERXYLARY PHLOEM (included phloem) By Marcelo R. Pace Interxylary phloem is the presence of phloem strands embedded within the secondary xylem (wood), and produced by the activity of a single cambium (Carlquist 2013). Stems with this cambial variant are also referred to as foraminate, due to the conspicuous interxylary phloem strands in the shape of dots scattered within the wood. (Fig. 1A). However, the presence of interxylary phloem is sometimes less evident and can only be confirmed by microscopy. Fig. 1. Stem cross-section of Strychnos (Loganiaceae). A. S. guianensis, macroscopic view. B. S. millepunctata, microscopic view. Interxylary phloem can have four different ontogenetic origins. The first one is where the cambium produces phloem in both directions (inside and outside), followed by the formation of xylem only towards the inside, and as a result, enclosing the phloem in the wood. Examples of this origin are present in Thunbergia (Acanthaceae; Fig. 2A) and Dicella (Malpighiaceae; Fig 2B). However, in Thunbergia the interxylary phloem is derived from the interfascicular cambium resulting in radial patches that alternate with regions of the xylem that originate from the fascicular cambium (Fig. 2A). A second origin of the interxylary phloem is through the formation of small phloem arcs. These later become embedded in the wood through the production of xylem by the cambium on their flanks. The resulting phloem islands will contain a fragment of cambium at the bottom. This type is present in Strychnos (Loganiaceae; Fig. 1), the African species of Combretum (Combretaceae; Van Vliet, 1979), and in at least one neotropical species of Combretum (Acevedo-Rodríguez, pers. -

Tissues and Other Levels of Organization MODULE - 1 Diversity and Evolution of Life
Tissues and Other Levels of Organization MODULE - 1 Diversity and Evolution of Life 5 Notes TISSUES AND OTHER LEVELS OF ORGANIZATION You have just learnt that cell is the fundamental structural and functional unit of organisms and that bodies of organisms are made up of cells of various shapes and sizes. Groups of similar cells aggregate to collectively perform a particular function. Such groups of cells are termed “tissues”. This lesson deals with the various kinds of tissues of plants and animals. OBJECTIVES After completing this lesson, you will be able to : z define tissues; z classify plant tissues; z name the various kinds of plant tissues; z enunciate the tunica corpus theory and histogen theory; z classify animal tissues; z describe the structure and function of various kinds of epithelial tissues; z describe the structure and function of various kinds of connective tissues; z describe the structure and function of muscular tissue; z describe the structure and function of nervous tissue. 5.1 WHAT IS A TISSUE Organs such as stem, and roots in plants, and stomach, heart and lungs in animals are made up of different kinds of tissues. A tissue is a group of cells with a common origin, structure and function. Their common origin means they are derived from the same layer (details in lesson No. 20) of cells in the embryo. Being of a common origin, there are similar in structure and hence perform the same function. Several types of tissues organise to form an organ. Example : Blood, bone, and cartilage are some examples of animal tissues whereas parenchyma, collenchyma, xylem and phloem are different tissues present in the plants. -

Tree Identification: from Bark and Leaves Or Needles
Tree Identification: from bark and leaves or needles A walk in the woods can be a lot of fun, especially if you bring your kids. How do you get them to come along with you? Tell them this. “Look at the bark on the trees. Can you can find any that look like burnt potato chips, warts, cat scratches, camouflage pants or rippling muscles?” Believe it or not, these are descriptions of different kinds of tree bark. Tree ID from bark:______________________________________________________ Black cherry: The bark looks like burnt potato chips. Hackberry: The bark is bumpy and warty. Ironwood: The bark has long thin strips. With a little imagination, an ironwood can look like it is used as a scratching post by cats. They can also be easily spotted in winter because their light brown dead leaves hang on well past the first snow. Sycamore: A tree with bark that looks like camouflaged pants. The largest tree in Illinois is a sycamore, a majestic 115-foot tree near Springfield. Musclewood: a tree with smooth gray bark covering a trunk with ridges that look like they are rippling muscles. Shagbark hickory: Long strips of shaggy bark peeling at both ends. Cottonwood: Has heavy ridges that make it look like Paul Bunyan’s corduroy pants. Bur oak: Thick and gnarly bark has deep craggy furrows. The corky bark allows it to easily withstand hot forest fires. Tree ID from leaves or needles:________________________________________ Willow Oak: Has elongated leaves similar to those of a willow tree. Red pine: Red has 3 letters; a red pine has groupings of 2-3 prickly needles. -
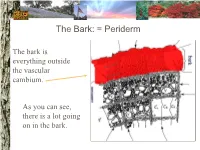
The Bark: = Periderm
The Bark: = Periderm The bark is everything outside the vascular cambium. As you can see, there is a lot going on in the bark. The Bark: periderm: phellogen (cork cambium): The phellogen is the region of cell division that forms the periderm tissues. Phellogen development influences bark appearance. The Bark: periderm: phellem (cork): Phellem replaces the epidermis as the tree increases in girth. Photosynthesis can take place in some trees both through the phellem and in fissures. The Bark: periderm: phelloderm: Phelloderm is active parenchyma tissue. Parenchyma cells can be used for storage, photosynthesis, defense, and even cell division! The Bark: phloem: Phloem tissue makes up the inner bark. However, it is vascular tissue formed from the vascular cambium. The Bark: phloem: sieve tube elements: Sieve tube elements actively transport photosynthates down the stem. Conifers have sieve cells instead. The cambium: The cambium is the primary meristem producing radial growth. It forms the phloem & xylem. The Xylem (wood): The xylem includes everything inside the vascular cambium. The Xylem: a growth increment (ring): The rings seen in many trees represent one growth increment. Growth rings provide the texture seen in wood. The Xylem: vessel elements: Hardwood species have vessel elements in addition to trachieds. Notice their location in the growth rings of this tree The Xylem: fibers: Fibers are cells with heavily lignified walls making them stiff. Many fibers in sapwood are alive at maturity and can be used for storage. The Xylem: axial parenchyma: Axial parenchyma is living tissue! Remember that parenchyma cells can be used for storage and cell division. -

Squamous Cell Carcinoma of the Renal Parenchyma
Zhang et al. BMC Urology (2020) 20:107 https://doi.org/10.1186/s12894-020-00676-5 CASE REPORT Open Access Squamous cell carcinoma of the renal parenchyma presenting as hydronephrosis: a case report and review of the recent literature Xirong Zhang1,2, Yuanfeng Zhang1, Chengguo Ge1, Junyong Zhang1 and Peihe Liang1* Abstract Background: Primary squamous cell carcinoma of the renal parenchyma is extremely rare, only 5 cases were reported. Case presentation: We probably report the fifth case of primary Squamous cell carcinoma (SCC) of the renal parenchyma in a 61-year-old female presenting with intermittent distending pain for 2 months. Contrast-enhanced computed tomography (CECT) revealed hydronephrosis of the right kidney, but a tumor cannot be excluded completely. Finally, nephrectomy was performed, and histological analysis determined that the diagnosis was kidney parenchyma squamous cell carcinoma involving perinephric adipose tissue. Conclusions: The present case emphasizes that it is difficult to make an accurate preoperative diagnosis with the presentation of hidden malignancy, such as hydronephrosis. Keywords: Kidney, Renal parenchyma, Squamous cell carcinoma, Hydronephrosis, Malignancy Background Case presentation Squamous cell carcinoma (SCC) of the renal pelvis is a The patient is a 61-year-old female. After suffering from rare neoplasm, accounting for only 0.5 to 0.8% of malig- intermittent pain in the right flank region for 2 months nant renal tumors [1], SCC of the renal parenchyma is she was referred to the urology department at an outside even less common. A review of the literature shows that hospital. The patient was diagnosed with hydronephrosis only five cases of primary SCC of the renal parenchyma of the right kidney and underwent a right ureteroscopy have been reported to date [2–6]. -

Secretory Tissues (Gesneriaceae)
Acta Bot. Neerl. 46(4), December 1997, p.413-420 Secretory tissues of the flower of Sanango racemosum (Gesneriaceae). I. Light microscopy Sara Maldonadoi* and Marisa Oteguiif * Institute de Recursos Bioldgicos, INTA 1712, Villa Udaondo, Castelar, Argentina; fFacultad de Ciencias La 1900 La Naturales y Museo, Universidad Nacional de Plata, Plata, Argentina SUMMARY Sanango racemosum (Ruiz & Pav.) Barringer has a dry stigma without a free-flowing secretion fluid but with a hydrated proteinaceous pellicle. The stigmatic surface is covered with unicellular, bottle-shaped papillae. At maturity, a viscous emulsion is accumulated between the cuticle and the pecto-cellulosic wall of the papillae, causing it to become detached from the surface of the papilla cell walls. The style has a central solid core of transmitting tissue. The cells of the transmitting tissue are rich in starch and exhibit thick lateral walls rich in pectic substance. The nectary disk is a ring elongated into a cup, with five lobes at the top. One of the most conspicuous histological features of the disk is the abundance of starch in the secretory cells. The disk is supplied only by phloem; the stomata are found in the top of the lobes. A fluid substance is produced just before anthesis and secreted through the stomata with no visible decline in starch level. During anthesis and after fertilization, a rapid decline in starch is observed. The hypothesis that the disk has other functions besides that of a nectary is discussed. Key-words: disk, nectary, osmophore, Sanango, stigma, transmitting tissue. INTRODUCTION The monotypic genus Sanango G. S. Bunting and J. -

Getting Chemicals Into Trees Without Spraying
Urban Forestry NR/FF/020 (pr) Getting Chemicals Into Trees Without Spraying Michael Kuhns, Forestry Extension Specialist This fact sheet provides an overview of injection, or phellogen that makes cork to thicken the outer bark, implantation, and other ways to get chemicals, mainly phloem that conducts food through the tree from where pesticides, into trees. Many techniques and systems it is stored or made to where it is being used (all of these exist and some are very good, some are good in some tissues together make up the bark), vascular cambium situations, and some are ineffective or bad for trees. that divides rapidly to make new phloem and xylem cells, This fact sheet addresses all of these. and xylem or wood. Xylem includes an outer layer called the sapwood that conducts mostly water and minerals from the roots to the canopy, and an inner layer called the Chemicals are applied to trees for many reasons. heartwood that is aged sapwood that has died and has lost Insecticides repel or kill damaging insects, fungicides its ability to conduct water but still adds strength. treat or prevent fungal diseases, nutrients and plant growth regulators affect growth, and herbicides kill trees or prevent sprouting after tree removal. Spraying is the most typical way to apply these chemicals. It is fast, uses readily available equipment, and is understood. The Phellem (Cork Phellogen down side of spraying is that much of the chemical being or Outer Bark) Phloem applied is wasted, either to drift, run off, or because it can not be applied precisely to where it is needed in the tree.