Boreal Plains Ecozone Evidence for Key Findings
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Information Package Watercourse
Information Package Watercourse Crossing Management Directive June 2019 Disclaimer The information contained in this information package is provided for general information only and is in no way legal advice. It is not a substitute for knowing the AER requirements contained in the applicable legislation, including directives and manuals and how they apply in your particular situation. You should consider obtaining independent legal and other professional advice to properly understand your options and obligations. Despite the care taken in preparing this information package, the AER makes no warranty, expressed or implied, and does not assume any legal liability or responsibility for the accuracy or completeness of the information provided. For the most up-to-date versions of the documents contained in the appendices, use the links provided throughout this document. Printed versions are uncontrolled. Revision History Name Date Changes Made Jody Foster enter a date. Finalized document. enter a date. enter a date. enter a date. enter a date. Alberta Energy Regulator | Information Package 1 Alberta Energy Regulator Content Watercourse Crossing Remediation Directive ......................................................................................... 4 Overview ................................................................................................................................................. 4 How the Program Works ....................................................................................................................... -

Transparency in Fisheries Management
Fish Habitat Restoration in Muriel Lake September 2018 Prepared by: Ray Makowecki MSc., BSc., BEd., P.Biol. 1 1975 2 3 PURPOSE • Muriel Lake Restoration • Do we have a Problem • Specific Fish and Fish Habitat Facts • Fish Habitat and Management Solutions 4 5 Muriel Lake 6800 ha at fsl 1981 - 559.1m Geodetic Elevation 1993 - 557.8m Geodetic Elevation 2013 - 555.7m Geodetic Elevation 2018 - 555.8m Geodetic Elevation 6 Muriel Lake 7m depth 2018 7 Lake Level 40 Year Trend of Declining Water Level • Current level • 555.8m 2018 FACTS & OBSERVATIONS What is the Problem? • Many older anglers indicate fishing was very good in the 1960’s, 1970’s and 1980’s when some whitefish, pike, perch and walleye would be caught and used. • Since 2000 little consumptive use 10 FACTS & OBSERVATIONS What is the Problem? •For over 20 years, several lakes in the area have had zero limits or very restrictive catch and keep limits. •Many lakes have due to low water levels been unable to support fish including Muriel. 11 FACTS & OBSERVATIONS What is the Problem? • Alberta’s commercial fishery was closed in 2014. Muriel Lake closed in 1996 • Anglers and fish consumers are not satisfied with prohibited consumptive use of Alberta’s publicly owned fish resources. 12 Annual Production - Muriel • Walleye 5848 kg/year (0.86 kg/ha) • Northern Pike 15,368 kg/year (2.26 kg/ha) • Yellow Perch 15,368 kg/year (2.26 kg/ha) • Lake Whitefish 29,920 (4.4 kg/ha) 13 Historic Harvest, Stocking and Regulations Muriel Lake Years Type of WALL NRPK YLPR LKWH Comments Fishery Commerc <20kg <1332 <10 <40,554 kg ial 1967-1976 Commerc Closed <223kg <1930 0 <25,890 ial 1996 1988-1996 Recreatio Catch 3 10 15 10 n limits Pre 1998 Recreatio Catch 0 0 0 0 n limits14 1998-2017 15 16 17 18 19 Muriel Lake 2012 • Government FWIN Testing 11 nets • Longnose Suckers (3) and Cyprinids (126) • No pike, whitefish, perch, burbot or walleye • Need some information here 20 Muriel Lake • Annual production for pike, lake whitefish and perch has not been harvested for the past 20 years. -

Village of Innisfree Audit Planning Report for the Year Ended December 31, 2019
Village of Innisfree Regular Council Meeting Innisfree Council Chambers, Innisfree AB December 17, 2019 @ 5:00 p.m. 1. Call to Order 2. Agenda a. Deletions/Additions b. Adoption of Agenda 3. Delegations 4. Adoption of Minutes a. November 19, 2019 Regular Council Minutes (Page 3-6) 5. Business Arising from the Minutes 6. Policies & Bylaws a. 645/19 – Amendment to Tax Rate Bylaw 637/19 (Page 7) 7. New Business a. 2020 Travel Lakeland Activity Guide (Page 8-14) b. 106.5 Country Christmas Advertising Packages (Page 15-16) c. ATB Financial – Loan Renewal Agreement (Page 17-19) d. Tax Forgiveness – Plan 4175R, Block 8, Lot 14 (Page 20-22) e. Alberta Municipal Affairs Letter December 4, 2019 – Municipal Accountability Program Review 2020 (Page 23-24) f. Federation of Canadian Municipalities Membership 2020-2021 (Page 25-26) g. Trans Canada Yellowhead Highway Association – Municipal Membership (Page 27-29) h. Notice of Hearing – Alberta Utilities Commission – Transmission Line 7L65 Rebuild Project (Page 30-33) i. Northern Lights Library System – 2020 Levy (Page 34) j. Metrix Group LLP - Engagement Letter & Audit Planning Report (Page 35-62) 8. Councillor Reports a. MMI FCSS Report – Deborah McMann (Page 63) b. Innisfree School Council Meeting – Deborah McMann (Page 64) 9. Administration Reports a. CAO Report & Action List (Page 65-68) b. Financials i. Tax Trial Balance – November 30, 2019 (Page 69) ii. Utility Trial Balance – November 30, 2019 (Page 70) iii. Accounts Payable – Cheque Listing – November, 2019 (Page 71-72) iv. Revenue & Expenses – November, 2019 (Page 73-78) c. Website Report (Page 79-80) 10. -

Jacobs Techmemo
450 - 1 Street SW Calgary, Alberta T2P 5H1 Tel: (403) 920-2940 Fax: (403) 920-2347 E-mail: [email protected] September 28, 2020 Filed Electronically Canada Energy Regulator Suite 210, 517 Tenth Avenue SW Calgary, Alberta T2R 0A8 Attention: Mr. Jean-Denis Charlebois, Secretary of the Commission Dear Mr. Charlebois: Re: NOVA Gas Transmission Ltd. (NGTL) 2018 Meter Stations and Laterals Abandonment Program (2018 Abandonment Program) MHW-003-2019, Order ZO-008-2019 and Amending Order AO-001-ZO-008-2019 (Order, as amended) Project Update: Saddle Lake West Lateral Abandonment File No.: OF-Fac-Gas-N081-2018-16 01 On December 20, 2019, the Canada Energy Regulator (CER or Commission) issued the Order, as amended, approving the 2018 Abandonment Program.1 NGTL writes to update the CER of a change in abandonment method for the Saddle Lake West Lateral, which was previously contemplated as part of the 2018 Abandonment Program application approved by the CER. Abandonment Method – Saddle Lake West Lateral In its application, NGTL proposed to abandon a 13.54 km section of the Saddle Lake West Lateral (SLW Lateral) located on and within Saddle Lake Cree Indian Reserve No. 125 (SLC IR No. 125) entirely by removal,2 and noted that while the lateral crosses two watercourses, being the Saddle Lake Creek and an unnamed tributary to the North Saskatchewan River (Watercourses), no in-stream or riparian work would be required.3 NGTL further advised it was working with Saddle Lake Cree Nation (SLCN) to conduct on-site field surveys to better understand site conditions for the SLW Lateral’s removal.4 NGTL has since conducted on-site field surveys in October 2018 and July 2020 with SLCN. -
Listing of Surface Water Quality Reports by Keywords
WATER POLICY BRANCH SURFACE WATER SECTION WATER QUALITY TEAM LISTING OF SURFACE WATER QUALITY REPORTS BY KEYWORDS HOW THIS LISTING WORKS: 1) Keywords are in alphabetical order. 2) Look up the Keyword in the Index; go to the listed page number. 3) The report title appears after the Keyword. A Report may appear twice, under two different Keywords. 4) Many reports are available electronically. Reports in .pdf format are hyperlinked (W####) from this Listing and may be downloaded here or through ESRD's Information Centre. Print copies can be obtained through the Information Centre or by e-mailing us at [email protected] 5) Some of the reports listed are available for loan through the Alberta Government Library. Contact them at [email protected] or (780) 427-2985. Updated: May 21, 2014 —A— Eutrophication ....................................................... 11 Agriculture .............................................................. 1 —F— Algae ....................................................................... 3 Field Lake ............................................................. 11 ALMS ..................................................................... 3 Fish ........................................................................ 11 AOSERP ................................................................. 1 Frank Lake ............................................................ 12 AOX ........................................................................ 1 —G— Aquatic Ecosystems ............................................... -

Glacitectonic Rafts and Their Role in the Generation of Quaternary Subglacial Bedforms and Deposits
Quaternary Research (2021), 1–35 doi:10.1017/qua.2021.11 Research Article Glacitectonic rafts and their role in the generation of Quaternary subglacial bedforms and deposits David J. A. Evansa*, Emrys R. Phillipsb and Nigel Atkinsonc aDepartment of Geography, Durham University, South Road, Durham DH1 3LE, UK; bBritish Geological Survey, Lyell Centre, Research Avenue South, Edinburgh EH14 4AP, UK and cAlberta Geological Survey, Twin Atria Building, Suite 402, 4999 98th Avenue, Edmonton, Alberta T6B 2X3, Canada Abstract Landforms and sediments on the palaeo–ice stream beds of central Alberta record glacitectonic raft production and subsequent progressive disaggregation and moulding, associated substrate ploughing, and grooving. We identify a subglacial temporal or developmental hierarchy that begins with incipient rafts, including en échelon hill-hole complexes, hill-hole pairs, and strike-slip raft complexes, all of which display patterns typical of transcurrent fault activation and pull apart. Many display jigsaw puzzle–style fragmentation, indicative of substrate displace- ment along shallow décollement zones and potentially related to patchy ice stream freeze-on. Their gradual fragmentation and smoothing pro- duces ice flow-transverse ridges (ribbed moraine), hill-groove pairs, and paraxial ridge and groove associations. Initiator scarp and megafluting associations are indicative of raft dislodgement and groove ploughing, leading to the formation of murdlins, crag-and-tails, stoss-and-lee type flutings and drumlins, and Type 1 hogsback flutings. Downflow modification of rafts creates linear block trains (rubble stripes), stoss-and-lee type megaflutings, horned crag-and-tails, rubble drumlinoids, and murdlins, diagnostic of an immature palaeo–ice stream footprint. Lateral ice stream margin migration ingests disaggregated thrust masses to form ridged spindles, ladder-type morphologies, and narrow zones of ribbed terrain and Type 2 hogsback flutings, an assemblage diagnostic of ice stream shear margin moraine formation. -

Soil Survey of the County of St. Paul, Alberta
Soi1 Survey of the County of St. Paul, Alberta J.A. Brierley, L.D. Andriashek and W.L. Nikiforuk Alberta Soi1 Survey Report No. 52 1993 - - - - - - - - - - - CENTRE OF LAND AND BIOLOGICAL RESOURCES RESEARCH CONTRIBUTION NO. 93-82 -_ PRINCIPALCOOPERATIVE AGENCIES: AGRICULTURE CANADA, CLBRR - ALBERTA RESEARCH COUNCIL, ENVIRONMENTAL RESEARCH AND ENGINEERING - - ‘- -- ii - - How to use this soi1 survey report - The soil survey report for the County of St. Paul consists Experience has shown that the following step by step of three chapters, five appendices and four 150 000 scale procedure assists users of the report: (lcm = 1/2km) planimetric base soi1 maps. A legend, 1. Locate the specific area on the appropiate map specific to each map sheet, describes the components of sheet. each soi1 map unit symbol. l an index to the four map sheets is on page 3 (a small The three chapters contain the following information: scale, index map is included on each map sheet) - Chapter 1 provides a general overview of the survey area l generally using the section, township and range grid, in terms of climatic, ecological and geologic charac- identii soi1 map delineation and the associated map teristics. symbol for the specific area. Chapter 2 provides a general description of the soi1 types 2. Expianation of map symbol. and associated landforms by subdividing the County into l find the map symbol and accompanying description of sixteen land systems. Each land system is described indi- associciated soi1 and landscape characteristics within vidually. the soi1 map unit legend, alphabetically and numerically Chapter 3 documents the methodology and assumptions arranged - used to rate the soi1 map units for arable agriculture and l soi1 map unit descriptions in the legend are described in - range productivity. -

The Buildingsmart Canada BIM Strategy
Leadership in Sustainable Infrastructure Leadership en Infrastructures Durables Vancouver, Canada May 31 – June 3, 2017/ Mai 31 – Juin 3, 2017 RECONSTRUCTION OF AREA-CAPACITY CURVES FOR ALBERTA LAKES Islam, Zahidul1, 3 and Seneka, Michael2 1, 2 Alberta Environment and Parks, Canada 3 [email protected] Abstract: Area-Capacity curves represent the relationship of surface area and volume of a lake or reservoir to elevation. Area-Capacity curves are essential to the development of water balance models for lakes; more specifically, they provide science-based information on surface water availability at any given time. This information can be vital to the execution of informed management decisions, such as the permitting of water withdrawals from lakes in such a way as to protect ecosystems while supporting reliable, quality water supplies that contribute to a sustainable economy. Currently, there are three major sources of area/capacity information in Alberta: the Atlas of Alberta Lakes (Mitchell 1990); Alberta Environment and Parks (AEP) archived files of historical bathymetric surveys and lake area/capacity calculations; and digital lake bathymetry from the Alberta Geological Survey (AGS). The Atlas of Alberta Lakes provides Area-Capacity curves for about 80 lakes in Alberta. However, these curves are only available on paper or in digital image format and are not usable unless digitized to retrieve the area/capacity information at various elevations. The AEP archived files contain manual calculations of area/capacity based on original bathymetry surveys for about 60 lakes and are not immediately accessible to the public. AGS data provide lakes bathymetry (GIS files) on their website, but not the actual information on area/capacity. -

NSWA Technical Bulletin Lake Level Trends in Alberta ‐ Preliminary Results
August 2017 www.nswa.ab.ca NSWA Technical Bulletin Lake Level Trends in Alberta ‐ Preliminary Results Chickakoo Lake – Bill Trout Introduction extremes). The Prairie provinces are particularly Canada is a land of lakes. It has more than a vulnerable to climate change effects, and some million lakes that cover 7.6% of the country’s area studies have described an overall pattern of th and is estimated to have approximately 20% of declining lake levels through most of the 20 4 the world’s existing freshwater reserves1. Lakes century . Some potential consequences of lake are an important source of water for a range of level fluctuations related to natural and human users (e.g. municipalities, agriculture, industry, stressors have been noticed in this region, for etc.), and are important ecosystems providing a example, water quality declines, increases in blue‐ multitude of ecological functions and a range of green algal warnings and fish kills. goods and recreational services. The North Saskatchewan Watershed Alliance Many studies in Canada have provided evidence (NSWA) encourages sustainable residential and of reduced water availability due to changing development practices in lake communities climate2,3. Lakes are not exempt from such within North Saskatchewan region. In this effects, and they are becoming more vulnerable context, the NSWA has been working with local to a range of stressors, both human (i.e. stewardship groups and municipalities to develop increasing consumptive use of water linked to lake management plans to protect and improve increased population and economic growth), and water quality and the health of the lakes. The natural (i.e. -
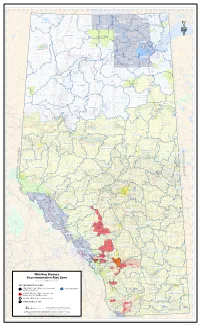
Whirling Disease Decontamination Risk Zone
TALTSON RIVER KAKISA RIVER ANDREW LAKE - TETHUL RIVER ANDREW LAKE - TETHUL RIVER MIDDLE SLAVE RIVER - SALT RIVER LITTLE BUFFALO RIVER PETITOT RIVER - BISTCHO LAKE WHITESAND - YATES RIVERS BUFFALO RIVER JACKFISH RIVER UPPER SLAVE RIVER LOWER HAY RIVER IN ALBERTA · NORTH LAKE ATHABASCA LOWER PEACE RIVER UPPER HAY RIVER IN ALBERTA WENTZEL RIVER OLD FORT RIVER - SOUTH LAKE ATHABASCA BOYER RIVER - PONTON RIVER McIVOR RIVER - LAKE CLAIRE LOWER BIRCH RIVER LOWER ATHABASCA RIVER FONTAS RIVER UPPER BIRCH RIVER KEG RIVER CHINCHAGA RIVER LOWER WABASCA RIVER MIKKWA RIVER SCULLY CREEK - PEACE RIVER ABOVE KEG RIVER FIREBAG RIVER BUFFALO - WOLVERINE RIVERS ELLS - STEEPBANK RIVERS NOTIKEWIN RIVER DOIG RIVER MACKAY RIVER UPPER WABASCA RIVER - PEERLESS LAKE CLEARWATER RIVER CADOTTE RIVER AND LAKE LOON RIVER - LUBICON RIVER HORSE RIVER CHURCHILL RIVER WHITEMUD RIVER - NORTH HEART RIVER CHURCHILL RIVER PEACE RIVER ABOVE PEACE RIVER CHRISTINA RIVER UTIKUMA RIVER - WABASCA LAKES PUSKWASKAU RIVER - LOWER SMOKY RIVER CALLING LAKE - HOUSE RIVER DILLON RIVER SOUTH HEART - EAST & WEST PRAIRIE RIVERS LOWER LITTLE SMOKY RIVER BEAVERLODGE RIVER NORTH LESSER SLAVE LAKE LOWER WAPITI RIVER - BEAR CREEK AND LAKE SAULTEAUX - DRIFTWOOD - LESSER SLAVE RIVERS WANDERING RIVER - LAC LA BICHE SAND RIVER MARTINEAU RIVER - COLD LAKE TAWATINAW RIVER UPPER WAPITI RIVER - NARRAWAY RIVER SIMONETTE RIVER UPPER BEAVER RIVER UPPER LITTLE SMOKY RIVER ATHABASCA BELOW FORT ASSINIBOINE KAKWA RIVER - CUTBANK RIVER FREEMAN RIVER LOWER PEMBINA RIVER JACKFISH - MURIEL CREEKS LOWER BEAVER RIVER