General Introduction
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pitt Water – Orielton Lagoon Tasmania Ecological Character
Pitt WaterWater – Orielton Lagoon Tasmania Ecological Character Description AugustDraft 1 2012August 2009 Pitt Water – Orielton Lagoon Tasmania Ecological Character Description August 2012 Introductory Notes The Environment Protection and Biodiversity Conservation Act 1999 (EPBC Act) prohibits actions that are likely to have a significant impact on the ecological character of a Ramsar wetland unless the Commonwealth Environment Minister has approved the taking of the action, or some other provision in the EPBC Act allows the action to be taken. The information in this ECD Publication does not indicate any commitment to a particular course of action, policy position or decision. Further, it does not provide assessment of any particular action within the meaning of the Environment Protection and Biodiversity Conservation Act 1999 (Cth), nor replace the role of the Minister or his delegate in making an informed decision to approve an action. The Water Act 2007 requires that in preparing the [Murray-Darling] Basin Plan, the Murray Darling Basin Authority (MDBA) must take into account Ecological Character Descriptions of declared Ramsar wetlands prepared in accordance with the National Framework. This ECD publication is provided without prejudice to any final decision by the Administrative Authority for Ramsar in Australia on change in ecological character in accordance with the requirements of Article 3.2 of the Ramsar Convention. Disclaimer While reasonable efforts have been made to ensure the contents of this ECD are correct, the Commonwealth of Australia as represented by the Department of Sustainability, Environment, Water, Population and Communities does not guarantee and accepts no legal liability whatsoever arising from or connected to the currency, accuracy, completeness, reliability or suitability of the information in this ECD. -

Evolutionary Patterns and Consequences of Developmental Mode in Cenozoic Gastropods from Southeastern Australia
Evolutionary patterns and consequences of developmental mode in Cenozoic gastropods from southeastern Australia Thesis submitted in accordance with the requirements of the University of Liverpool for the degree of Doctor in Philosophy by Kirstie Rae Thomson September 2013 ABSTRACT Gastropods, like many other marine invertebrates undergo a two-stage life cycle. As the adult body plan results in narrow environmental tolerances and restricted mobility, the optimum opportunity for dispersal occurs during the initial larval phase. Dispersal is considered to be a major influence on the evolutionary trends of different larval strategies. Three larval strategies are recognised in this research: planktotrophy, lecithotrophy and direct development. Planktotrophic larvae are able to feed and swim in the plankton resulting in the greatest dispersal potential. Lecithotrophic larvae have a reduced planktic period and are considered to have more restricted dispersal. The planktic period is absent in direct developing larvae and therefore dispersal potential in these taxa is extremely limited. Each of these larval strategies can be confidently inferred from the shells of fossil gastropods and the evolutionary trends associated with modes of development can be examined using both phylogenetic and non-phylogenetic techniques. This research uses Cenozoic gastropods from southeastern Australia to examine evolutionary trends associated with larval mode. To ensure the species used in analyses are distinct and correctly assigned, a taxonomic review of the six families included in this study was undertaken. The families included in this study were the Volutidae, Nassariidae, Raphitomidae, Borsoniidae, Mangeliidae and Turridae. Phylogenetic analyses were used to examine the relationships between taxa and to determine the order and timing of changes in larval mode throughout the Cenozoic. -

THE BIRDS of IRELAND. Order PASSERES. Family TURDIDAE
THE BIRDS OF IRELAND. Order PASSERES. Family TURDIDAE Subfamily TURDINNAE THE MISTLE-THRUSH. Turdus viscivorus, Linnæus. Though now resident, common, and widely distributed, this bird was apparently unknown in Ireland before the nineteenth century. A Mistle-Thrush, shot early in 1808 in the Co. Antrim was the first Irish example that Thompson had heard of during the first half of the century the spread and increase of this species was noticed throughout Ireland, the Western coast being apparently the last reached. Thus Mr. W. Sinclair remembers it in Tyrone in1820, but it arrived in Western Donegal ten years later. Up to 1860 it was unknown in Achill, but now breeds there. In Western Connaught it is scarce, but is believed still to be on the increase throughout Ireland in general. Flocks occur from June to the end of autumn, and may be seen in the same locality for days together. These perform local migrations, but disperse before winter. In November there is an immigration from Great Britain, though on a much smaller scale than that of the Song-Thrush or Blackbird. Some Mistle-Thrushes visit Rathlin Island in frosts, and Valentia is another winter resort. The Mistle-Thrush is a welcome addition to the song-birds of Ireland, from October onwards it may be heard in our mild climate; at first rarely, but more frequently and fully as each winter month passes, until the full song is delivered in February and March; while even frost and snow will not always silence it. I have heard one uttering its notes in short snatches between each triple stroke of its wings while it flew. -

Pitt Water – Orielton Lagoon Tasmania Ecological Character Description Augustdraft 1 August2012 2009
Pitt Water – Orielton Lagoon Tasmania Ecological Character Description AugustDraft 1 August2012 2009 Pitt Water – Orielton Lagoon Tasmania Ecological Character Description August 2012 Introductory Notes The Environment Protection and Biodiversity Conservation Act 1999 (EPBC Act) prohibits actions that are likely to have a significant impact on the ecological character of a Ramsar wetland unless the Commonwealth Environment Minister has approved the taking of the action, or some other provision in the EPBC Act allows the action to be taken. The information in this ECD Publication does not indicate any commitment to a particular course of action, policy position or decision. Further, it does not provide assessment of any particular action within the meaning of the Environment Protection and Biodiversity Conservation Act 1999 (Cth), nor replace the role of the Minister or his delegate in making an informed decision to approve an action. The Water Act 2007 requires that in preparing the [Murray-Darling] Basin Plan, the Murray Darling Basin Authority (MDBA) must take into account Ecological Character Descriptions of declared Ramsar wetlands prepared in accordance with the National Framework. This ECD publication is provided without prejudice to any final decision by the Administrative Authority for Ramsar in Australia on change in ecological character in accordance with the requirements of Article 3.2 of the Ramsar Convention. Disclaimer While reasonable efforts have been made to ensure the contents of this ECD are correct, the Commonwealth of Australia as represented by the Department of Sustainability, Environment, Water, Population and Communities does not guarantee and accepts no legal liability whatsoever arising from or connected to the currency, accuracy, completeness, reliability or suitability of the information in this ECD. -

The Significance of Rubbish Tips As an Additional Food Source for the Kelp Gull and the Pacific Gull in Tasmania
The Significance of Rubbish Tips as an Additional Food Source for the Kelp Gull and the Pacific Gull in Tasmania by e. e--~'" (& G. M. Coulson, B.A. (Hans.), Dip.Ed. (Melb.) and .J~ R.I. Coulson, B .Sc., Dip.Ed. (Melb.) Being a thesis submitted in part fulfilment of the requirements for the degree of Master of Environmental Studies Centre for Environmental Studies University of Tasmania August, 1982 nl\, (l,_ '/ Cl-- \'\~'.2. ACKNOWLEDGEMENTS We are grateful to our supervisors, Dr. A.M.M. Richardson and Dr. J.J. Todd, and our external advisors, Mr. A.W.J. Fletcher and Mr. J.G.K. Harris, for their guidance in the preparation and planning of this thesis. Mrs. L. Ramsay typed the thesis. Mr. D. Barker (Tasmanian Museum, Hobart) and Mr. R.H. Green (Queen Victoria Museum, Launceston) gave us access to their gull collections. Mr. G. Davis assisted with the identification of chitons. Officers of the Tasmanian National Parks and Wildlife Service provided information, assistance, equipment and specimens, and officers of the Tasmanian Department of the Environment gave us information on tips. The cities and municipalities in south-east Tasmania granted permission to study gulls at tips under their control, and we are particularly grateful for the co-operation shown by council staff at Clarence, Hobart and Kingborough tips. Mr. R. Clark allowed us to observe gulls at Richardson's Meat Works, Lutana. CONTENTS ABSTRACT 1 1. INTRODUcriON 3 2. GULL POPULATIONS IN THE NORTHERN HEMISPHERE 7 2.1 Population Increases 8 2.1.1 Population size and rates·of change 10 2.1.2 Growth of breeding colonies 13 2.2 Reasons for the Population Increase 18 2.2.1 Protection 18 2.2.2 Greater food availability 19 2.3 Effects of Population Increase 22 2.3.1 Competition with other species 22 2.3.2 Agricultural pests 23 2.3.3 Public health risks 24 2.3.4 Urban nesting 25 2.3.5 Aircraft bird-strikes 25 2.4 Future Trends 26 3. -

The First Australian Breeding Record of the White-Fronted Tern* by J
VOL. 8 (5) MARCH, 1980 137 The First Australian Breeding Record of the White-fronted Tern* By J. S. WHINRAY, Flinders Island, Tasmania, 7255. Early in 1979 I found some White-fronted Terns Sterna striata Gmelin breeding in Tasmania on Battery Island in the Furneaux Group. Pre· viously the species was known to breed only in New Zealand (Hindwood, 1946; Falla, et al., 1966; Serventy, et al., 1971 ; Frith, 1977). Hindwood (1946) brought together most of the prior records of this species for the Australian region and, with few exceptions, these fell within the period of May to November. Hitchcock & Favaloro (1951) considered it "more than likely" that some of Hindwood's late Sydney records, particularly the January "stragglers", were referable to the Com mon Tern Sterna hirundo longipennis. The subsequent Australian records of the White-fronted Tern also fell within the May to November period (the sources include Hitchcock & Favaloro, 1951; Clark & Dawson, 1957; Lawrence, 1959; and Kinsky, 1963). The New Zealand breeding season starts in October (Oliver, 1955; Falla, et al., 1966). So the period of the species' absence from the Aus tralian region coincided with much of the New Zealand breeding season. The first six Australian recoveries of birds banded in New Zealand were made in 1955 in New South Wales. Six sightings of banded birds were made the next year in the same state. These records were " the first definite evidence to support the theory that White-fronted Terns winter ing on the coast of New South Wales and Victoria are birds of New Zealand origin" (Clarke & Dawson, 1957). -
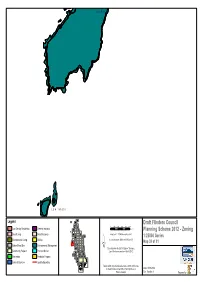
Draft Flinders Council Planning Scheme 2012
Joins Map 34 L o w I s l e t s Legend Draft Flinders Council 0 200 400 600 800 1,000 Low Density Residential General Industrial Metres Planning Scheme 2012 - Zoning Rural Living Rural Resource Map Scale 1 : 25,000 when printed at A3 1:25000 Series Environmental Living Utilities Coordinate System: GDA 1994 MGA Zone 55 Map 39 of 81 Urban Mixed Use Environmental Management ´ Base data from theLIST, © State of Tasmania Community Purpose Port and Marine Land title data current as of 30/04/2012 Recreation Particular Purpose General Business Locality Boundary Before taking any action based on data shown on this map it should first be verified with a Planning Officer of Date: 15/05/2012 Flinders Council. Doc. Version: 0 Prepared by L o Joins Map 35 n g g P o i n t t R o a d Whitemark Chalky Island Joins Map 41 Reef Island Mile Island Little Chalky Island R e e f I s l a n d Isabella Island Legend Draft Flinders Council 0 200 400 600 800 1,000 Low Density Residential General Industrial Metres Planning Scheme 2012 - Zoning Rural Living Rural Resource Map Scale 1 : 25,000 when printed at A3 1:25000 Series Environmental Living Utilities Coordinate System: GDA 1994 MGA Zone 55 Map 40 of 81 Urban Mixed Use Environmental Management ´ Base data from theLIST, © State of Tasmania Community Purpose Port and Marine Land title data current as of 30/04/2012 Recreation Particular Purpose General Business Locality Boundary Before taking any action based on data shown on this map it should first be verified with a Planning Officer of Date: 15/05/2012 Flinders Council. -

The Little Grassbird in the Furneaux Group, Tasmania. by J
Dec. ] WHINRAY: Little Grassbird in Furneaux Group 321 1976 The Little Grassbird in the Furneaux Group, Tasmania. By J. S. WHINRAY, Flinders Idand, Tasmania. On the Tasmanian mainland the Little Grassbird Megalurus gramineus occurs in "marshes, water-filled ditches and reedy channels" (Sharland, 1958). On King Island, in western Bass Strait, it has been recorded recently in swampy areas (Green and McGarvie, 1971). The recent records for Flinders Island, in the Furneaux Group in south-eastern Bass Strait, were made in the "thickly growing reeds of two lagoons" (Green, 1969) . However the Little Grassbird has been recorded for three small Furneaux Group islands but no comment has yet been made on the difference between this habitat and the usual Tasmanian habitat (White, 1909; Cashion, 1958; Green, 1969; Newman, 1974). Recently I have found Little Grassbirds on seven more small Furneaux Group islands. Here are brief descriptions of these ten islands and notes on the occurrence of the species. Except for Samphire Island, the descriptions are drawn from my unpublished field notes. Cat Island, about 30 Ha. in area, has a main vegetation of tussock grass which is mainly Poa poiformis but includes a broken coastal belt of Stipa teretifolia. There is one tiny patch of sand dune shrubs including Myoporum insulare and Olearia axillaris (both to 2.2m high). There are also two tiny patches of shrubby Atriplex cinerea (to 1.2 m). It is a waterless island. In December 1908 Little Grassbirds "were flushed now and again from the tall grass" (White, 1909) and Cashion recorded them "frequently" between mid January and mid March, 1957 (Cashion, 1958). -

BIC-1967.Pdf
TABLE OF CONTENTS Page Secretaries' Report ... ... ... ... 3 Treasurer's Report ... ... ... ... 4 Subscriptions, etc. ... ... ... ... ... 9 Obituaries ... ... ... ... ... 9 Predannack Nature Reserve ... ... ... 9 List of Contributors ... ... ... ... 10 Cornish Records ... ... ... ... ... 11 Corrections to 1966 Report ... ... ... ... 48 Cornish Seas ... ... ... ... ... 49 Ringing Recoveries ... ... ... 50 Isles of Scilly Records ... ... ... ... 51 Round Island ... ... ... ... ... 67 Porthgwarra Report ... ... ... ... 68 Breeding Birds of the Bodmin area ... ... 71 The Torrey Canyon Incident and Sea Birds of Cornwall 78 After the Torrey Canyon ... ... ... ... 90 Committees ... ... ... ••• ••• 130 List of Members ... ... ... ... ••• 131 Rules 147 1 THIRTY-SEVENTH REPORT OF The Cornwall Bird-Watching and Preservation Society 1967 Edited by J. E. BECKERLEGGE assisted by J. B. and S. BOTTOMLEY, A. G. PARSONS and N. R. PHILLIPS SECRETARIES' REPORT FOR 1967 On December 31st the Membership of the Society was 866. 112 new Members were elected during the year and 35 Members were lost by resignations and death, giving a net gain of 77. Two General Meetings, three Executive Committee Meetings and 10 Field Days were held. At the Annual General Meeting Mr. N. Dymond, of the British Trust for Ornithology, spoke on the work of a Bird ringing station and illustrated his talk with colour slides. At the Autumn Meeting Mr. T. O. Darke showed a film illustrating Bird life throughout the year in the Porthcothan area, near Padstow. During the year the following species of special interest were noted: White-billed Diver, Wilson's Phalarope, Long-billed Dowitcher, Roller, Orphean Warbler, Parula Warbler and American Redstart: this last being the first record for Britain. Records for Cornwall, arranged in the order of this Report, should be sent to the Reverend J. -

2ND EDITION OILE Á in the IRISH ISLANDS GUIDE Oileáin Noun Islands (Irish) the IRISH ISLANDS GUIDE
OILEÁIN 2ND EDITION OILE Á THE IRISH ISLANDS GUIDE IN Oileáin noun Islands (Irish) THE IRISH ISLANDS GUIDE OVER 570 IRISH ISLANDS DAVID WALSH A wealth of information on the wildlife, stories and history of the Irish islands. David Walsh For those wishiing to visit in small boats or kayaks there are details of: OILEÁIN Landings • ISBN 978-1-906095-37-6 ND DITION 10000 2 E • Camping • Drinking water • Tidal information 9781906 095376 The Irish islands guide Other Guides from Pesda Press The North West The North East The East Coast The Mid West The South Coast www.pesdapress.com DAVID WALSH OILEÁIN 2ND EDITION The Irish islands guide Second Edition 2014 First Published in Great Britain 2004 by Pesda Press Unit 22, Galeri Doc Victoria Caernarfon Gwynedd LL55 1SQ © Copyright 2004 Dave Walsh ISBN: 978−1−906095−37−6 The Author asserts the moral right to be identified as the author of this work. All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, electronic, mechanical, photocopying, recording or otherwise, without the prior written permission of the Publisher. Printed and bound in Poland. www.Pozkal.com 2 Introduction Oileáin is a detailed guide to almost every Irish coastal island. The guide is comprehensive, describing over 570 islands and islets, big and small, far out to sea and close in by the shore, inhabited and uninhabited, worth the trouble visiting or not. Oileáin has always told it as it is, reef by reef, rock by rock, good or bad, pleasant or otherwise, but now even the more so. -

The Lighthouse Work of Sir James T. Chance
THE LIGHTHOUSE WORK OF SIR JAMES CHANCE, BARONET THE LIGHTHOUSE WORK OF SIR JAMES CHANCE BARONET BY JAMES FREDERICK CHANCE, M.A. WITH A PREFACE BY JAMES KENWARD, C.E., F.S.A. LONDON SMITH, ELDER, & CO., 15 WATERLOO PLACE 1902 [All rights reserved] BIOGRAPHICAL NOTE JAMES TIMMINS CHANCE was born on March 22, 1814, being the oldest son of Mr. William Chance, of Birmingham. From an early age he showed evidence of unusual talent, studying with success not only mathematics and natural science, in which subjects he gained high honours at the London University (now University College), but also classics and modern languages, and even Hebrew. At the age of nineteen he proceeded to Trinity College, Cambridge, and graduated in 1838 as Seventh Wrangler. He also began the study of law, entering as a student of Lincoln's Inn in 1836. But circumstances obliged him, immediately on leaving Cambridge, to enter the glass- making firm of Chance Brothers & Co., in which his father was a partner, and he remained himself a partner therein for fifty years, being head of it for twenty-five. Apart from this work, he interested himself greatly in social questions, particularly in the promotion of education; and he was a liberal and constant donor in a great many directions, his two principal benefactions being the gift and endow- ment of a public park at West Smethwick and the foundation of the ‘Chance School of Engineering’ at Birmingham University. He was for many years a director of the London and North-Western Railway, Served the office of High Sheriff of Staffordshire in 1868, and in later years was a valued member of the Council of University College. -
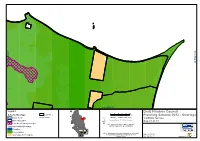
Draft Flinders Council Planning Scheme 2012
Joins Map 28 Joins Map 26 Joins Map 33 Legend Draft Flinders Council 0 200 400 600 800 1,000 Scheme Overlays Land Titles Metres Planning Scheme 2012 - Overlays Airport Buffer Roads Map Scale 1 : 25,000 when printed at A3 1:25000 Series Coordinate System: GDA 1994 MGA Zone 55 Buffer/Attenuation Map 27 of 81 Local Scenic Management Area Base data from theLIST, © State of Tasmania Regional Model Overlays Land title data current as of 30/04/2012 Flood Risk Priority Habitat Before taking any action based on data shown on this map Acid Sulfate Soil Probability it should first be verified with a Planning Officer of Date: 16/05/2012 Flinders Council. Doc. Version: 0 Prepared by Babel Island Joins Map 27 C a t I s l a n d Storehouse Island Legend Draft Flinders Council 0 200 400 600 800 1,000 Scheme Overlays Land Titles Metres Planning Scheme 2012 - Overlays Airport Buffer Roads Map Scale 1 : 25,000 when printed at A3 1:25000 Series Coordinate System: GDA 1994 MGA Zone 55 Buffer/Attenuation Map 28 of 81 Local Scenic Management Area Base data from theLIST, © State of Tasmania Regional Model Overlays Land title data current as of 30/04/2012 Flood Risk Priority Habitat Before taking any action based on data shown on this map Acid Sulfate Soil Probability it should first be verified with a Planning Officer of Date: 16/05/2012 Flinders Council. Doc. Version: 0 Prepared by Joins Map 30 Wybalenna Island Prime Seal Island Joins Map 34 Legend Draft Flinders Council 0 200 400 600 800 1,000 Scheme Overlays Land Titles Metres Planning Scheme 2012 - Overlays Airport Buffer Roads Map Scale 1 : 25,000 when printed at A3 1:25000 Series Coordinate System: GDA 1994 MGA Zone 55 Buffer/Attenuation Map 29 of 81 Local Scenic Management Area Base data from theLIST, © State of Tasmania Regional Model Overlays Land title data current as of 30/04/2012 Flood Risk Priority Habitat Before taking any action based on data shown on this map Acid Sulfate Soil Probability it should first be verified with a Planning Officer of Date: 16/05/2012 Flinders Council.