<I>Heteroconger Digueti</I>
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pacific Plate Biogeography, with Special Reference to Shorefishes
Pacific Plate Biogeography, with Special Reference to Shorefishes VICTOR G. SPRINGER m SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY • NUMBER 367 SERIES PUBLICATIONS OF THE SMITHSONIAN INSTITUTION Emphasis upon publication as a means of "diffusing knowledge" was expressed by the first Secretary of the Smithsonian. In his formal plan for the Institution, Joseph Henry outlined a program that included the following statement: "It is proposed to publish a series of reports, giving an account of the new discoveries in science, and of the changes made from year to year in all branches of knowledge." This theme of basic research has been adhered to through the years by thousands of titles issued in series publications under the Smithsonian imprint, commencing with Smithsonian Contributions to Knowledge in 1848 and continuing with the following active series: Smithsonian Contributions to Anthropology Smithsonian Contributions to Astrophysics Smithsonian Contributions to Botany Smithsonian Contributions to the Earth Sciences Smithsonian Contributions to the Marine Sciences Smithsonian Contributions to Paleobiology Smithsonian Contributions to Zoo/ogy Smithsonian Studies in Air and Space Smithsonian Studies in History and Technology In these series, the Institution publishes small papers and full-scale monographs that report the research and collections of its various museums and bureaux or of professional colleagues in the world cf science and scholarship. The publications are distributed by mailing lists to libraries, universities, and similar institutions throughout the world. Papers or monographs submitted for series publication are received by the Smithsonian Institution Press, subject to its own review for format and style, only through departments of the various Smithsonian museums or bureaux, where the manuscripts are given substantive review. -

Updated Checklist of Marine Fishes (Chordata: Craniata) from Portugal and the Proposed Extension of the Portuguese Continental Shelf
European Journal of Taxonomy 73: 1-73 ISSN 2118-9773 http://dx.doi.org/10.5852/ejt.2014.73 www.europeanjournaloftaxonomy.eu 2014 · Carneiro M. et al. This work is licensed under a Creative Commons Attribution 3.0 License. Monograph urn:lsid:zoobank.org:pub:9A5F217D-8E7B-448A-9CAB-2CCC9CC6F857 Updated checklist of marine fishes (Chordata: Craniata) from Portugal and the proposed extension of the Portuguese continental shelf Miguel CARNEIRO1,5, Rogélia MARTINS2,6, Monica LANDI*,3,7 & Filipe O. COSTA4,8 1,2 DIV-RP (Modelling and Management Fishery Resources Division), Instituto Português do Mar e da Atmosfera, Av. Brasilia 1449-006 Lisboa, Portugal. E-mail: [email protected], [email protected] 3,4 CBMA (Centre of Molecular and Environmental Biology), Department of Biology, University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal. E-mail: [email protected], [email protected] * corresponding author: [email protected] 5 urn:lsid:zoobank.org:author:90A98A50-327E-4648-9DCE-75709C7A2472 6 urn:lsid:zoobank.org:author:1EB6DE00-9E91-407C-B7C4-34F31F29FD88 7 urn:lsid:zoobank.org:author:6D3AC760-77F2-4CFA-B5C7-665CB07F4CEB 8 urn:lsid:zoobank.org:author:48E53CF3-71C8-403C-BECD-10B20B3C15B4 Abstract. The study of the Portuguese marine ichthyofauna has a long historical tradition, rooted back in the 18th Century. Here we present an annotated checklist of the marine fishes from Portuguese waters, including the area encompassed by the proposed extension of the Portuguese continental shelf and the Economic Exclusive Zone (EEZ). The list is based on historical literature records and taxon occurrence data obtained from natural history collections, together with new revisions and occurrences. -

FISHES (C) Val Kells–November, 2019
VAL KELLS Marine Science Illustration 4257 Ballards Mill Road - Free Union - VA - 22940 www.valkellsillustration.com [email protected] STOCK ILLUSTRATION LIST FRESHWATER and SALTWATER FISHES (c) Val Kells–November, 2019 Eastern Atlantic and Gulf of Mexico: brackish and saltwater fishes Subject to change. New illustrations added weekly. Atlantic hagfish, Myxine glutinosa Sea lamprey, Petromyzon marinus Deepwater chimaera, Hydrolagus affinis Atlantic spearnose chimaera, Rhinochimaera atlantica Nurse shark, Ginglymostoma cirratum Whale shark, Rhincodon typus Sand tiger, Carcharias taurus Ragged-tooth shark, Odontaspis ferox Crocodile Shark, Pseudocarcharias kamoharai Thresher shark, Alopias vulpinus Bigeye thresher, Alopias superciliosus Basking shark, Cetorhinus maximus White shark, Carcharodon carcharias Shortfin mako, Isurus oxyrinchus Longfin mako, Isurus paucus Porbeagle, Lamna nasus Freckled Shark, Scyliorhinus haeckelii Marbled catshark, Galeus arae Chain dogfish, Scyliorhinus retifer Smooth dogfish, Mustelus canis Smalleye Smoothhound, Mustelus higmani Dwarf Smoothhound, Mustelus minicanis Florida smoothhound, Mustelus norrisi Gulf Smoothhound, Mustelus sinusmexicanus Blacknose shark, Carcharhinus acronotus Bignose shark, Carcharhinus altimus Narrowtooth Shark, Carcharhinus brachyurus Spinner shark, Carcharhinus brevipinna Silky shark, Carcharhinus faiformis Finetooth shark, Carcharhinus isodon Galapagos Shark, Carcharhinus galapagensis Bull shark, Carcharinus leucus Blacktip shark, Carcharhinus limbatus Oceanic whitetip shark, -

Contents of Volume 42 (2012)
CONTENTS OF VOLUME 42 (2012) Issue 1 Tripp-Valdez A., Arreguín-Sánchez F., Zetina-Rejón M.J. The food of Selene peruviana (Actinopterygii: Perciformes: Carangidae) in the southern Gulf of California .................. 1 Zarrad R., Alemany F., Jarboui O., Garcia A., Akrout F. Comparative characterization of the spawning environments of European anchovy, Engraulis encrasicolus , and round sardinella, Sardinella aurita (Actinopterygii: Clupeiformes) in the eastern coast of Tunisia ............................... 9 Ordines F., Valls M., Gouraguine A. Biology, feeding, and habitat preferences of Cadenat’s rockfish, Scorpaena loppei (Actinopterygii: Scorpaeniformes: Scorpaenidae), in the Balearic Islands (western Mediterranean) ..................................... 21 Socha M., Sokołowska-Mikołajczyk M., Szczerbik P., Chyb J., Mikołajczyk T., Epler P. The effect of polychlorinated biphenyls mixture (Aroclor 1254) on the embryonic development and hatching of Prussian carp, Carassius gibelio , and common carp, Cyprinus carpio (Actinopterygii: Cypriniformes: Cyprinidae) ............................................................................................................................................................... 31 Khan A., Ghosh K. Characterization and identification of gut-associated phytase-producing bacteria in some fresh water fish cultured in ponds ...................................................................................................................................................................................... -

Training Manual Series No.15/2018
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by CMFRI Digital Repository DBTR-H D Indian Council of Agricultural Research Ministry of Science and Technology Central Marine Fisheries Research Institute Department of Biotechnology CMFRI Training Manual Series No.15/2018 Training Manual In the frame work of the project: DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals 2015-18 Training Manual In the frame work of the project: DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals 2015-18 Training Manual This is a limited edition of the CMFRI Training Manual provided to participants of the “DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals” organized by the Marine Biotechnology Division of Central Marine Fisheries Research Institute (CMFRI), from 2nd February 2015 - 31st March 2018. Principal Investigator Dr. P. Vijayagopal Compiled & Edited by Dr. P. Vijayagopal Dr. Reynold Peter Assisted by Aditya Prabhakar Swetha Dhamodharan P V ISBN 978-93-82263-24-1 CMFRI Training Manual Series No.15/2018 Published by Dr A Gopalakrishnan Director, Central Marine Fisheries Research Institute (ICAR-CMFRI) Central Marine Fisheries Research Institute PB.No:1603, Ernakulam North P.O, Kochi-682018, India. 2 Foreword Central Marine Fisheries Research Institute (CMFRI), Kochi along with CIFE, Mumbai and CIFA, Bhubaneswar within the Indian Council of Agricultural Research (ICAR) and Department of Biotechnology of Government of India organized a series of training programs entitled “DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals”. -

Reef Fishes of the Bird's Head Peninsula, West Papua, Indonesia
Check List 5(3): 587–628, 2009. ISSN: 1809-127X LISTS OF SPECIES Reef fishes of the Bird’s Head Peninsula, West Papua, Indonesia Gerald R. Allen 1 Mark V. Erdmann 2 1 Department of Aquatic Zoology, Western Australian Museum. Locked Bag 49, Welshpool DC, Perth, Western Australia 6986. E-mail: [email protected] 2 Conservation International Indonesia Marine Program. Jl. Dr. Muwardi No. 17, Renon, Denpasar 80235 Indonesia. Abstract A checklist of shallow (to 60 m depth) reef fishes is provided for the Bird’s Head Peninsula region of West Papua, Indonesia. The area, which occupies the extreme western end of New Guinea, contains the world’s most diverse assemblage of coral reef fishes. The current checklist, which includes both historical records and recent survey results, includes 1,511 species in 451 genera and 111 families. Respective species totals for the three main coral reef areas – Raja Ampat Islands, Fakfak-Kaimana coast, and Cenderawasih Bay – are 1320, 995, and 877. In addition to its extraordinary species diversity, the region exhibits a remarkable level of endemism considering its relatively small area. A total of 26 species in 14 families are currently considered to be confined to the region. Introduction and finally a complex geologic past highlighted The region consisting of eastern Indonesia, East by shifting island arcs, oceanic plate collisions, Timor, Sabah, Philippines, Papua New Guinea, and widely fluctuating sea levels (Polhemus and the Solomon Islands is the global centre of 2007). reef fish diversity (Allen 2008). Approximately 2,460 species or 60 percent of the entire reef fish The Bird’s Head Peninsula and surrounding fauna of the Indo-West Pacific inhabits this waters has attracted the attention of naturalists and region, which is commonly referred to as the scientists ever since it was first visited by Coral Triangle (CT). -

Anguilliformes, Saccopharyngiformes, and Notacanthiformes (Teleostei: Elopomorpha)
* Catalog of Type Specimens of Recent Fishes in the National Museum of Natural History, Smithsonian Institution, 6: Anguilliformes, Saccopharyngiformes, and Notacanthiformes (Teleostei: Elopomorpha) DAVID G. SMITH I SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY • NUMBER 566 SERIES PUBLICATIONS OF THE SMITHSONIAN INSTITUTION Emphasis upon publication as a means of "diffusing knowledge" was expressed by the first Secretary of the Smithsonian. In his formal plan for the Institution, Joseph Henry outlined a program that included the following statement: "It is proposed to publish a series of reports, giving an account of the new discoveries in science, and of the changes made from year to year in all branches of knowledge." This theme of basic research has been adhered to through the years by thousands of titles issued in series publications under the Smithsonian imprint, commencing with Smithsonian Contributions to Knowledge in 1848 and continuing with the following active series: Smithsonian Contributions to Anthropology Smithsonian Contributions to Astrophysics Smithsonian Contributions to Botany Smithsonian Contributions to the Earth Sciences Smithsonian Contributions to the Marine Sciences Smithsonian Contributions to Paleobiology Smithsonian Contributions to Zoology Smithsonian Folklife Studies Smithsonian Studies in Air and Space Smithsonian Studies in History and Technology In these series, the Institution publishes small papers and full-scale monographs that report the research and collections of its various museums and bureaux or of professional colleagues in the world of science and scholarship. The publications are distributed by mailing lists to libraries, universities, and similar institutions throughout the world. Papers or monographs'submitted for series publication are received by the Smithsonian Institution Press, subject to its own review for format and style, only through departments of the various Smithsonian museums or bureaux, where the manuscripts are given substantive review. -

Natural History Observations of Hawaiian Garden Eels, Gorgasia Hawaiiensis (Congridae: Heterocongrinae), from the Island of Hawai’I
Natural history observations of Hawaiian garden eels, Gorgasia hawaiiensis (Congridae: Heterocongrinae), from the Island of Hawai’i By Emily Donham*, Michael S. Foster, Marc R. Rice, Gregor M. Cailliet, Mary M. Yoklavich, and Scott L. Hamilton Abstract Garden eels occur worldwide in the tropics, but little is known about their biology and ecology. We studied Hawaiian garden eel (Gorgasia hawaiiensis) colonies near Kawaihae, Hawai’i to investigate multiple aspects of the basic biology of this species. Colonies of G. hawaiiensis occurred at depths from 16 to 36 m in soft bottom habitat adjacent to rocky reefs. Highest burrow densities (up to 40 eels m-2) were in shallower water, and large (~10 mm diameter) burrows were more abundant, less dense and commonly found in pairs in deeper water. Eels emerged around sunrise, and withdrew and covered burrow entrances around sunset. Age was estimated from annual rings in sectioned otoliths (n = 17), and modeled to suggest fast growth to a maximum size of ~600 mm total length and a maximum age of six years. Prey size and eel anatomy suggest these fish feed by ingesting planktonic prey and processing them in the esophagus. The most common food items were small (<0.5 mm) demersal harpacticoid, cyclopoid and calanoid copepods, and unidentified fish eggs. These and other observations indicate that G. hawaiiensis is abundant, has a high population turnover rate, and may enrich sandy bottom habitat within their beds by facilitating energy flow from the water column to the benthos. *Corresponding Author E-mail: [email protected] Pacific Science, vol. 71, no. -

Spotted Garden
Spotted Garden Eel • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • Heteroconger hassi osa Classificationthrough What human understanding groups does this organism belong to based on characteristics shared with other organisms? Class: Actinopterygii (ray-finned fishes) Order: Aulopiformes (lizardfishes and relatives) Family: Congridae (marine eels) Genus and Species: Heteroconger hassi (spotted garden eel) Distribution Where in the world does this species live? Spotted garden eels live in the warm parts of the Pacific Ocean and the Red Sea. Habitat: What kinds of areas does this species live in? They live on the sandy flats that border coral reefs at depths of 23’ – 150’ (7–45m). They can also be found in areas dense with seagrass. They dig a burrow into the sandy sea bottom using their pointed tail. When the burrow is deep enough they use their dorsal fin to push the sand out of the burrow. Slime that they secrete from their skin cements the burrow walls so that the burrow will not collapse. Physical Description How would this animal’s body shape and size be described? • They resemble a worm. • They grow up to 16” (40cm) long • Their body diameter is .5” (1.3cm). • Adults have a white body covered in small black spots with three large black patches on their body. • Juveniles are entirely black. • They have large yellow eyes. • They have an upturned mouth. • They are sexually dimorphic, meaning males and females look different. The male is much larger than the female and it’s jaw sticks out further. Diet What does this species eat? In their historic range: They eat tiny planktonic animals that float by them in the water current. -
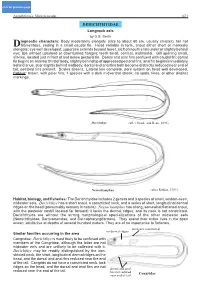
DERICHTHYIDAE Longneck Eels by D.G
click for previous page Anguilliformes: Muraenesocidae 1671 DERICHTHYIDAE Longneck eels by D.G. Smith iagnostic characters: Body moderately elongate (size to about 60 cm, usually smaller), tail not Dfilamentous, ending in a small caudal fin. Head variable in form, snout either short or markedly elongate; eye well developed; upper jaw extends beyond lower, cleft of mouth ends under or slightly behind eye; lips without upturned or downturned flanges; teeth small, conical, multiserial. Gill opening small, slit-like, located just in front of and below pectoral fin. Dorsal and anal fins confluent with caudal fin; dorsal fin begins on anterior third of body, slightly behind tip of appressed pectoral fins; anal fin begins immediately behind anus, at or slightly behind midbody; dorsal and anal fins both become distinctly reduced near end of tail; pectoral fins present. Scales absent. Lateral line complete, pore system on head well developed. Colour: brown, with paler fins; 1 species with a dark midventral streak; no spots, lines, or other distinct markings. Derichthys (after Goode and Bean, 1896) Nessorhamphus (after Robins, 1989) Habitat, biology, and fisheries: The Derichthyidae includes 2 genera and 3 species of small, seldom-seen, midwater eels. Derichthys has a short snout, a constricted neck, and a series of short, longitudinal dermal ridges on the head (presumably sensory in nature). Nessorhamphus has a long, somewhat flattened snout, with the posterior nostril located far forward; it lacks the dermal ridges, and its neck is not constricted. Derichthyids are without the strong morphological specializations of the other midwater eels (Nemichthyidae, Serrivomeridae, and Saccopharyngiformes). They spend their entire lives in the open ocean; adults live at depths of several hundred meters. -

Norntates PUBLISHED by the AMERICAN MUSEUM of NATURAL HISTORY CENTRAL PARK WEST at 79TH STREET, NEW YORK, N.Y
AMERICAN MUSEUM Norntates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N.Y. 10024 Number 3037, 13 pp., 5 figures, 3 tables February 26, 1992 Systematic Significance of the Burrow Form of Seven Species of Garden Eels (Congridae: Heterocongnnae) JAMES C. TYLER' AND C. LAVETT SMITH2 ABSTRACT Garden eels are highly specialized anguilliform tude and wavelength in relation to the length of fishes that live in colonies worldwide in warm seas. the fish and the size of the casts have potential Unlike most eels, they are sight feeders, spending significance in classification. their daylight hours partially extended from their A preliminary analysis using Hennig86 indi- burrows feeding on drifting particles. Because they cates that the two species of Gorgasia in our sam- live in unconsolidated sand it has been difficult to ple are indeed each other's closest relatives and determine the form oftheir burrow. Using araldite part of a lineage that includes Heteroconger klau- epoxy plastic, we made casts ofthe burrows of six sewitzi and H. hassi. The relationships ofH. halis, species representing both of the currently recog- H. perissodon, and H. polyzona, however, cannot nized genera. All of the casts are in the form of be resolved from our data. simple sinusoidal waves but differences in ampli- INTRODUCTION Congrid eels of the subfamily Heterocon- in the eastern Pacific, eastern Atlantic, and grinae are ecologically notable because all of western Atlantic (personal commun. of P. the species live in burrows in colonies of up Castle and J. Randall, who are revising the to around 10,000 diurnal, planktivorous in- subfamily). -

Species Acromycter Nezumi (Asano, 1958)
FAMILY Congridae Kaup, 1856 - conger eels SUBFAMILY Congrinae Kaup, 1856 - conger eels [=Atteridi, Ophioides, Leptocephalini, Congeridae, Macrocephenchelyidae, Scalanogoinae, Urocongrinae] GENUS Acromycter Smith & Kanazawa, 1977 - conger eels Species Acromycter alcocki (Gilbert & Cramer, 1897) - Alcock's congrid eel Species Acromycter atlanticus Smith, 1989 - Atlantic conger eel Species Acromycter longipectoralis Karmovskaya, 2004 - longfinned conger eel Species Acromycter nezumi (Asano, 1958) - Nezumi's conger eel Species Acromycter perturbator (Parr, 1932) - perturbator conger eel GENUS Bassanago Whitley, 1948 - conger eels Species Bassanago albescens (Barnard, 1923) - hairy conger Species Bassanago bulbiceps Whitley, 1948 - swollen-headed conger eel Species Bassanago hirsutus (Castle, 1960) - deepsea conger Species Bassanago nielseni (Karmovskaya, 1990) - Nielsen's conger eel GENUS Bathycongrus Ogilby, 1898 - conger eels [=Congrina, Microcephalocongrus, Pseudoxenomystax, Rhechias, Uranoconger] Species Bathycongrus aequoreus (Gilbert & Cramer, 1897) - Kaiwi bathycongrus [=armiger] Species Bathycongrus bertini (Poll, 1953) - Poll's conger Species Bathycongrus bleekeri Fowler, 1934 - Bleeker's bathycongrus Species Bathycongrus bullisi (Smith & Kanazawa, 1977) - bullish conger Species Bathycongrus dubius (Breder, 1927) - dubius conger Species Bathycongrus guttulatus (Günther, 1887) - lined conger [=roosendaali, stimpsoni] Species Bathycongrus longicavis Karmovskaya, 2009 - Vanuatu conger Species Bathycongrus macrocercus (Alcock, 1894) -