Inca Project No
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
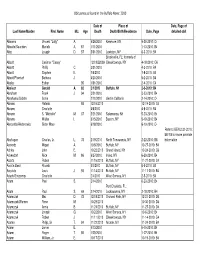
Obituaries Buffalo News 2010 by Name
Obituaries as found in the Buffalo News: 2010 Date of Place of Date, Page of Last Name/Maiden First Name M.I. Age Death Death/Birth/Residence Date, Page detailed obit Abbarno Vincent "Lolly" A. 9/26/2010 Kenmore, NY 9-30-2010: C4 Abbatte/Saunders Murielle A. 87 1/11/2010 1-13-2010: B4 Abbo Joseph D. 57 5/31/2010 Lewiston, NY 6-3-2010: B4 Brooksville, FL; formerly of Abbott Casimer "Casey" 12/19/22009 Cheektowaga, NY 4-18-2010: C6 Abbott Phillip C. 3/31/2010 4-3-2010: B4 Abbott Stephen E. 7/6/2010 7-8-2010: B4 Abbott/Pfoetsch Barbara J. 4/20/2010 5-2-2010: B4 Abeles Esther 95 1/31/2010 2-4-2010: C4 Abelson Gerald A. 82 2/1/2010 Buffalo, NY 2-3-2010: B4 Abraham Frank J. 94 3/21/2010 3-23-2010: B4 Abrahams/Gichtin Sonia 2/10/2010 died in California 2-14-2010: C4 Abramo Rafeala 93 12/16/2010 12-19-2010: C4 Abrams Charlotte 4/6/2010 4-8-2010: B4 Abrams S. "Michelle" M. 37 5/21/2010 Salamanca, NY 5-23-2010: B4 Abrams Walter I. 5/15/2010 Basom, NY 5-19-2010: B4 Abrosette/Aksterowicz Sister Mary 6/18/2010 6-19-2010: C4 Refer to BEN 2-21-2010: B6/7/8 for more possible Abshagen Charles, Jr. L. 73 2/19/2010 North Tonawanda, NY 2-22-2010: B8 information Acevedo Miguel A. 10/6/2010 Buffalo, NY 10-27-2010: B4 Achkar John E. -

The German Surname Atlas Project ± Computer-Based Surname Geography Kathrin Dräger Mirjam Schmuck Germany
Kathrin Dräger, Mirjam Schmuck, Germany 319 The German Surname Atlas Project ± Computer-Based Surname Geography Kathrin Dräger Mirjam Schmuck Germany Abstract The German Surname Atlas (Deutscher Familiennamenatlas, DFA) project is presented below. The surname maps are based on German fixed network telephone lines (in 2005) with German postal districts as graticules. In our project, we use this data to explore the areal variation in lexical (e.g., Schröder/Schneider µtailor¶) as well as phonological (e.g., Hauser/Häuser/Heuser) and morphological (e.g., patronyms such as Petersen/Peters/Peter) aspects of German surnames. German surnames emerged quite early on and preserve linguistic material which is up to 900 years old. This enables us to draw conclusions from today¶s areal distribution, e.g., on medieval dialect variation, writing traditions and cultural life. Containing not only German surnames but also foreign names, our huge database opens up possibilities for new areas of research, such as surnames and migration. Due to the close contact with Slavonic languages (original Slavonic population in the east, former eastern territories, migration), original Slavonic surnames make up the largest part of the foreign names (e.g., ±ski 16,386 types/293,474 tokens). Various adaptations from Slavonic to German and vice versa occurred. These included graphical (e.g., Dobschinski < Dobrzynski) as well as morphological adaptations (hybrid forms: e.g., Fuhrmanski) and folk-etymological reinterpretations (e.g., Rehsack < Czech Reåak). *** 1. The German surname system In the German speech area, people generally started to use an addition to their given names from the eleventh to the sixteenth century, some even later. -

Синергистички Ефекат Блокаде Il-33/St2 И Pdl/Pd-1 Осовина На Прогресију Мишјег Карцинома Дојке
УНИВЕРЗИТЕТ У КРАГУЈЕВЦУ ФАКУЛТЕТ МЕДИЦИНСКИХ НАУКА Марина Јовановић Синергистички ефекат блокаде IL-33/ST2 и PDL/PD-1 осовина на прогресију мишјег карцинома дојке ДОКТОРСКА ДИСЕРТАЦИЈА КРАГУЈЕВАЦ, 2021. UNIVERSITY OF KRAGUJEVAC FACULTY OF MEDICAL SCIENCES Marina Jovanovic Synergistical effect of IL-33/ST2 and PDL/PD-1 blockage in a mammary carcinoma Doctoral Dissertation KRAGUJEVAC, 2021. Идентификациона страница докторске дисертације Аутор Име и презиме: Марина Јовановић Датум и место рођења: 21.11.1991. Ћуприја Садашње запослење: Клинички центар Крагујевац, специјализант оториноларингологије Докторска дисертација Наслов: Синергистички ефекат блокаде IL-33/ST2 и PDL/PD-1 осовина на прогресију мишјег карцинома дојке Број страница: 138 Број слика: 55 (53 графикона, 2 табеле) Број библиографских података: 312 Установа и место где је рад израђен: Факултет медицинских наука у Крагујевцу Научна област (УДК): Медицина Ментор: Проф. др Иван Јовановић, ванредни професор Факултета медицинских наука Универзитета у Крагујевцу за уже научне области Микробиологија и имунологија, Онкологија Оцена и одбрана Датум пријаве теме: Број одлуке и датум прихватања теме докторске/уметничке дисертације: IV-03-65/21 од 04.09.2019. године Комисија за оцену научне заснованости теме и испуњености услова кандидата: 1. Проф. др Небојша Арсенијевић, редовни професор Факултета медицинских наука Универзитета у Крагујевцу за уже научне области Микробиологија и имунологија; Онкологијa, председник 2. Проф. др Гордана Радосављевић, ванредни професор Факултета медицинских наука Универзитета у Крагујевцу за ужу научну област Микробиологија и имунологија, члан 3. Доц. др Милан Јовановић, доцент Медицинског факултета Војномедицинске академије Универзитета одбране у Београду за ужу научну област Хирургија, члан Комисија за оцену и одбрану докторске/уметничке дисертације: Датум одбране дисертације: 1 Doctoral dissertation identification page Author Name and surname: Marina Jovanovic Date and place of birth: 21.11.1991. -

EJTS European Journal of Transformation Studies 2020, Vol.8, Supplement 1
EJTS European Journal of Transformation Studies 2020, Vol.8, Supplement 1 1 EJTS European Journal of Transformation Studies 2020, Vol.8, Supplement 1 EUROPEAN JOURNAL OF TRANSFORMATION STUDIES 2020 Vol. 8 Supplement 1 © by Europe Our House, Tbilisi e-ISSN 2298-0997 2 EJTS European Journal of Transformation Studies 2020, Vol.8, Supplement 1 Arkadiusz Modrzejewski University of Gdansk, Poland [email protected] Editors Tamar Gamkrelidze Europe Our House, Tbilisi, Georgia Tatiana Tökölyová Ss. Cyril and Methodius University in Trnava, Slovakia Rafał Raczyński Research Institute for European Policy, Poland Paweł Nieczuja–Ostrowski – executive editor Pomeranian University in Slupsk, Poland Jaroslav Mihálik – deputy editor Ss. Cyril and Methodius University in Trnava, Slovakia Edita Poórová – copy editor Ss. Cyril and Methodius University in Trnava, Slovakia Andrii Kutsyk– assistant editor Lesya Ukrainka Eastern European National University, Lutsk, Ukraine Editorial Advisory Board Prof. Jakub Potulski,University of Gdansk, Poland – chairperson Prof. Tadeusz Dmochowski,University of Gdansk, Poland Prof. Slavomír Gálik, University of Ss.Cyril and Methodius in Trnava, Slovakia Prof. Wojciech Forysinski, Eastern Mediterranean University, Famangusta, Northern Cyprus Prof. Danuta Plecka, Zielona Gora University, Poland Prof. Anatoliy Kruglashov, Chernivtsi National University, Ukraine Prof. Malkhaz Matsaberidze, Ivane Javakashvili Tbilisi State University Prof. Ruizan Mekvabidze, Gori State Teaching University, Georgia Prof. Lucia Mokrá, Comenius University in Bratislava, Slovakia Prof. Andras Bozoki, Central European University in Budapest, Hungary Prof. Tereza - Brînduşa Palade, National University of Political and Public Administra- tion in Bucharest, Romania Prof. Elif Çolakoğlu, Atatürk University in Erzurum, Turkey Prof. Valeriu Mosneaga, Moldova State University in Chişinău, Republic of Moldova Prof. Andrei Taranu, National University of Political Science and Public Administration in Bucharest, Romania Prof. -

Roman Catholic Missionaries and La Mission Ambulante with the Métis, Plains Cree and Blackfoot, 1840-1880
Les missionnaires sauvages: Roman Catholic missionaries and la mission ambulante with the Métis, Plains Cree and Blackfoot, 1840-1880 Mario Giguère Department of History McGill University, Montréal Submitted August 2009 A thesis submitted to McGill University in partial fulfillment of the requirements of the degree of M.A. in History. This thesis is copyright © 2009 Mario Giguère Library and Archives Bibliothèque et Canada Archives Canada Published Heritage Direction du Branch Patrimoine de l’édition 395 Wellington Street 395, rue Wellington Ottawa ON K1A 0N4 Ottawa ON K1A 0N4 Canada Canada Your file Votre référence ISBN: 978-0-494-66115-4 Our file Notre référence ISBN: 978-0-494-66115-4 NOTICE: AVIS: The author has granted a non- L’auteur a accordé une licence non exclusive exclusive license allowing Library and permettant à la Bibliothèque et Archives Archives Canada to reproduce, Canada de reproduire, publier, archiver, publish, archive, preserve, conserve, sauvegarder, conserver, transmettre au public communicate to the public by par télécommunication ou par l’Internet, prêter, telecommunication or on the Internet, distribuer et vendre des thèses partout dans le loan, distribute and sell theses monde, à des fins commerciales ou autres, sur worldwide, for commercial or non- support microforme, papier, électronique et/ou commercial purposes, in microform, autres formats. paper, electronic and/or any other formats. The author retains copyright L’auteur conserve la propriété du droit d’auteur ownership and moral rights in this et des droits moraux qui protège cette thèse. Ni thesis. Neither the thesis nor la thèse ni des extraits substantiels de celle-ci substantial extracts from it may be ne doivent être imprimés ou autrement printed or otherwise reproduced reproduits sans son autorisation. -

Delinquent Current Year Real Property
Delinquent Current Year Real Property Tax as of February 1, 2021 PRIMARY OWNER SECONDARY OWNER PARCEL ID TOTAL DUE SITUS ADDRESS 11 WESTVIEW LLC 964972494700000 1,550.02 11 WESTVIEW RD ASHEVILLE NC 1115 INVESTMENTS LLC 962826247600000 1,784.57 424 DEAVERVIEW RD ASHEVILLE NC 120 BROADWAY STREET LLC 061935493200000 630.62 99999 BROADWAY ST BLACK MOUNTAIN NC 13:22 LEGACIES LLC 967741958700000 2,609.06 48 WESTSIDE VILLAGE RD UNINCORPORATED 131 BROADWAY LLC 061935599200000 2,856.73 131 BROADWAY ST BLACK MOUNTAIN NC 1430 MERRIMON AVENUE LLC 973095178600000 2,759.07 1430 MERRIMON AVE ASHEVILLE NC 146 ROBERTS LLC 964807218300000 19,180.16 146 ROBERTS ST ASHEVILLE NC 146 ROBERTS LLC 964806195600000 17.24 179 ROBERTS ST ASHEVILLE NC 161 LOGAN LLC 964784681600000 1,447.39 617 BROOKSHIRE ST ASHEVILLE NC 18 BRENNAN BROKE ME LLC 962964621500000 2,410.41 18 BRENNAN BROOK DR UNINCORPORATED 180 HOLDINGS LLC 963816782800000 12.94 99999 MAURICET LN ASHEVILLE NC 233 RIVERSIDE LLC 963889237500000 17,355.27 350 RIVERSIDE DR ASHEVILLE NC 27 DEER RUN DRIVE LLC 965505559900000 2,393.79 27 DEER RUN DR ASHEVILLE NC 28 HUNTER DRIVE REVOCABLE TRUST 962421184100000 478.17 28 HUNTER DR UNINCORPORATED 29 PAGE AVE LLC 964930087300000 12,618.97 29 PAGE AVE ASHEVILLE NC 299 OLD HIGHWAY 20 LLC 971182306200000 2,670.65 17 STONE OWL TRL UNINCORPORATED 2M HOME INVESTMENTS LLC 970141443400000 881.74 71 GRAY FOX DR UNINCORPORATED 311 ASHEVILLE CONDO LLC 9648623059C0311 2,608.52 311 BOWLING PARK RD ASHEVILLE NC 325 HAYWOOD CHECK THE DEED! LLC 963864649400000 2,288.38 325 HAYWOOD -

Senate the Senate Met at 3 P.M
E PL UR UM IB N U U S Congressional Record United States th of America PROCEEDINGS AND DEBATES OF THE 116 CONGRESS, FIRST SESSION Vol. 165 WASHINGTON, MONDAY, SEPTEMBER 9, 2019 No. 143 Senate The Senate met at 3 p.m. and was Washington is where we come to work. of nominations to important Federal called to order by the President pro We come here to fight for our neigh- offices. The American people deserve to tempore (Mr. GRASSLEY). bors and for the places we love and are be governed by the government they f proud to hail from. voted for, and every time we confirm The American people know this is a another one of the uncontroversial, PRAYER highly charged political moment. They amply qualified public servants whom The Chaplain, Dr. Barry C. Black, of- haven’t sent us here to stage pitched President Trump has selected for these fered the following prayer: battles or score political points. They executive branch posts, we fulfill a Let us pray. elected us to make a difference for constitutional responsibility and make Almighty God, sovereign of our Na- them and their families. We do that by manifest the people’s decision. tion and Lord of our lives, thank You taking care of the people’s business and Of course, in the days and weeks for infusing us with the confidence that by collaborating in good faith to com- ahead, another major duty before us You order our steps each day. plete our work and attend to the press- will be the appropriations process. -

Final Report Ica Project No. 223 the Biological
FINAL REPORT ICA PROJECT NO. 223 THE BIOLOGICAL IMPORTANCE OF COPPER A Literature Review June, 1995 The contractor who produced this report is an independent contractor and is not an agent of ICA. ICA makes no express or implied warranty with regard to the information contained in this report. ICA PROJECT 223 Preface In 1973 the International Copper Research Association Incorporated initiated a grant to review the literature dealing with the biological importance of copper in marine and estuarine environments. This was followed by a second review in 1978. It was then apparent that a very large number of publications concerning copper in the marine environment were appearing each year and that an annual review was appropriate. Reviews prior to 1984 considered copper only in marine and estuarine environments. However, events occurring on land and in freshwater were often mentioned because chemical and biological factors and processes pertinent to one environment could often be applied to the others. As a result, the review became larger, covering not only freshwater, saltwater and terrestrial environments but also agriculture and medicine. It was apparent from the literature that most of the general concepts about the importance and the effects of copper could be applied in all environments. This also meant that an understanding of the environmental chemistry of copper could be applied in medicine as well as agriculture, the marine environment as well as soils. The reviews pointed out the broad application of concepts about the biological importance as well as the environmental chemistry of copper. The present review includes literature for the period 1992-1993. -

Aes Corporation
THE AES CORPORATION THE AES CORPORATION The global power company A Passion to Serve A Passion A PASSION to SERVE 2000 ANNUAL REPORT ANNUAL REPORT THE AES CORPORATION 1001 North 19th Street 2000 Arlington, Virginia 22209 USA (703) 522-1315 CONTENTS OFFICES 1 AES at a Glance AES CORPORATION AES HORIZONS THINK AES (CORPORATE OFFICE) Richmond, United Kingdom Arlington, Virginia 2 Note from the Chairman 1001 North 19th Street AES OASIS AES TRANSPOWER Arlington, Virginia 22209 Suite 802, 8th Floor #16-05 Six Battery Road 5 Our Annual Letter USA City Tower 2 049909 Singapore Phone: (703) 522-1315 Sheikh Zayed Road Phone: 65-533-0515 17 AES Worldwide Overview Fax: (703) 528-4510 P.O. Box 62843 Fax: 65-535-7287 AES AMERICAS Dubai, United Arab Emirates 33 AES People Arlington, Virginia Phone: 97-14-332-9699 REGISTRAR AND Fax: 97-14-332-6787 TRANSFER AGENT: 83 2000 AES Financial Review AES ANDES FIRST CHICAGO TRUST AES ORIENT Avenida del Libertador COMPANY OF NEW YORK, 26/F. Entertainment Building 602 13th Floor A DIVISION OF EQUISERVE 30 Queen’s Road Central 1001 Capital Federal P.O. Box 2500 Hong Kong Buenos Aires, Argentina Jersey City, New Jersey 07303 Phone: 852-2842-5111 Phone: 54-11-4816-1502 USA Fax: 852-2530-1673 Fax: 54-11-4816-6605 Shareholder Relations AES AURORA AES PACIFIC Phone: (800) 519-3111 100 Pine Street Arlington, Virginia STOCK LISTING: Suite 3300 NYSE Symbol: AES AES ENTERPRISE San Francisco, California 94111 Investor Relations Contact: Arlington, Virginia USA $217 $31 Kenneth R. Woodcock 93% 92% AES ELECTRIC Phone: (415) 395-7899 $1.46* 91% Senior Vice President 89% Burleigh House Fax: (415) 395-7891 88% 1001 North 19th Street $.96* 18 Parkshot $.84* AES SÃO PAULO Arlington, Virginia 22209 Richmond TW9 2RG $21 Av. -

1986 Surname
Surname Given Age Date Page Maiden Note Abegg Missel 88 15-Dec C-8 Abernathy Manuel 79 1-Jan C-5 Abner Tom 71 24-Jun B-7 Abraham Aloysius J. 77 24-Jul C-2 Abram Harold Glenn 75 14-Sep D-2 Abramson Frances L. 46 28-Dec C-7 Levine Ackerman Mary 79 16-Sep B-7 Adam Paul T. 76 10-Sep C-5 Adams Claude 78 20-Jan A-5 Adams Gloria L. 48 23-Sep D-6 Adams Irene 77 21-Aug C-3 Adams Martha 72 3-Nov C-1 Adank Gerald C. 65 1-Dec C-8 Veteran of World War II Adkins Johnnie Lee 78 13-Feb B-9 Adley Daisy A. 93 2-Sep D-7 Ahlborn Raymond W. 73 2-May C-1 Aird Gordon R. 77 21-Nov D-1 See article, p. D-1 Aitken Marion 70 1-Jul B-7 Aksentijevic Martha 56 17-Sep B-8 Alamillo Nora 75 4-Feb C-1 Albert Lester E. 65 30-Sep B-7 Albrecht Victoria A. 84 3-Jan A-7 Aldrin Raymond E. 71 11-Aug B-5 Aleksandrovic Ivan 76 11-Dec B-13 Aleksandrovic Jelena 79 2-Dec C-1 Ales Francis J. 69 31-Mar B-8 Alexander Janet 49 28-Jul C-1 Alexander Penny C. 60 24-Mar C-1 Alexander Terry 53 9-Dec C-1 Alexander Vera (Cook) 2-Dec C-1 Alexander William A. 50 27-Dec C-2 Alfaro Mark A. 21 5-Feb D-1 Alger Kenneth H. -

Graduatoria Provvisoria C.S
FASCI UFFICIO CODICE CODICE LINGUA CODICE COGNOME NOME POSIZIO INCLUS PUNTI TOTALI PREFEREN NUMER SERVIZI ISTITUTO(PRE ISTITUTO O CON O SENZA PROVINC PROFIL FERENZA DI (PRESENTAZI RISERV DEMERIT A IALE SEDE) O ONE NE A ZE O FIGLI O 3 RM RMIC8D500D CS I RMTF200009DOMANDA) ACCHIONI TIZIANA 1 41,03 17; 3 RM RMIC8D500D CS I RMTD48000N ORLANDO RACHELE 2 40,12 18; 1 3 RM RMIC8D500D CS I RMIC8AY002 PENNELLA MARIA LUIGIA 3 39,85 17; X 3 RM RMIC8D500D CS I RMIC8BA001 DE ROSA CARMINA 4 39,30 3 RM RMIC8D500D CS I RMIS05200R PLACENTINO GIULIA 5 39,15 17; 18; 1 3 RM RMIC8D500D CS I RMSL04000R ROSSETTI MONIA 6 38,50 18; 1 3 RM RMIC8D500D CS I RMIC8BG00X FRISON MARZIA 7 35,90 17; 18; 3 3 RM RMIC8D500D CS I RMIC8D500D LORENA RAFFAELE 8 35 18; 2 3 RM RMIC8D500D CS I RMPC29000G PETRILLO CRESCENZO 9 35 3 RM RMIC8D500D CS I RMIC8F600E CANTELLI GABRIELE 10 34,80 18; 1 X 3 RM RMIC8D500D CS I RMEE30700B MICOZZI FRANCA 11 30,50 17; X 3 RM RMIC8D500D CS I RMIC8BF004 MEO GAVINO 12 30,30 17; 18; 1 3 RM RMIC8D500D CS I RMRI08000G OFFREDA LUCA 13 30,30 3 RM RMIC8D500D CS I RMIS05200R STAMPA ELEONORA 14 30,25 17; 18; 2 X 3 RM RMIC8D500D CS I RMIC8EW00X ROBUSTELLI LUCIO 15 29,70 3 RM RMIC8D500D CS I RMIC81500N DE PIERRO MADDALENA 16 29,07 17; 3 RM RMIC8D500D CS I RMIC8FT003 SAVINO SONIA 17 28,85 3 RM RMIC8D500D CS I RMIC8FT003 CARRATU' PASQUALINO 18 28,80 19; 3 RM RMIC8D500D CS I RMIC8BH00Q PIGLIUCCI VIVIANA 19 28,25 3 RM RMIC8D500D CS I RMIC8F9002 DE LUZI KATIUSCIA 20 27,71 17; 18; 1 X 3 RM RMIC8D500D CS I RMIS02400L AIELLO IDA 21 27,64 17; 18; 2 3 RM RMIC8D500D CS -

International Handbook of Occupational Therapy Interventions Ingrid Söderback Editor
International Handbook of Occupational Therapy Interventions Ingrid Söderback Editor International Handbook of Occupational Therapy Interventions Editor Ingrid Söderback Associate Professor in Occupational Therapy and Rehabilitation Karolinska Institute, Stockholm, Sweden and University Lecturer Emerita Department of Public Health and Caring Sciences Uppsala University, Uppsala, Sweden Home address: Sickla Kanalgata 31 nb SE 13165 Nacka E-mail: [email protected] ISBN: 978-0-387-75423-9 e-ISBN: 978-0-387-75424-6 DOI: 10.1007/978-0-387-75424-6 Springer Dordrecht Heidelberg London New York Library of Congress Control Number: 2009927464 © Springer Science+Business Media, LLC 2009 All rights reserved. This work may not be translated or copied in whole or in part without the written permission of the publisher (Springer Science+Business Media, LLC, 233 Spring Street, New York, NY 10013, USA), except for brief excerpts in connection with reviews or scholarly analysis. Use in connection with any form of information storage and retrieval, electronic adaptation, computer software, or by similar or dissimilar methodology now known or hereafter developed is forbidden. The use in this publication of trade names, trademarks, service marks, and similar terms, even if they are not identified as such, is not to be taken as an expression of opinion as to whether or not they are subject to proprietary rights. Printed on acid-free paper Springer is part of Springer Science+Business Media (www.springer.com) To be occupied is a fundamental right in every human being’s life. The occupational therapists’ main professional role is to encourage the clients’ occupational performances. To occupational therapy students, occupational therapists, and members of rehabilitation and health care teams, who are professionally working to improve their clients’ health and wellness, and to stakeholders responsible for the administration of occupational therapy, worldwide.