Changes in Forest Structure in the Bavarian Forest National Park – an Evaluation After 10 Years of the BIOKLIM-Project
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Potential European Bison (Bison Bonasus) Habitat in Germany
PROJECT REPORT Potential European bison (Bison bonasus) habitat in Germany Tobias Kuemmerle Humboldt-University Berlin Benjamin Bleyhl Humboldt-University Berlin Wanda Olech University of Warsaw & European Bison Friends Society Kajetan Perzanowski Carpathian Wildlife Research Station MIZ, PAS & European Bison Friends Society 1 PROJECT REPORT CONTENTS CONTENTS ...............................................................................................................................................................2 INTRODUCTION .......................................................................................................................................................3 HISTORIC DISTRIBUTION OF EUROPEAN BISON IN EUROPE .....................................................................................4 EUROPEAN BISON HABITAT PREFERENCES ..............................................................................................................6 HABITAT USE OF CONTEMPORARY EUROPEAN BISON POPULATIONS .......................................................................................... 6 PALEO-ECOLOGICAL DATA ON EUROPEAN BISON HABITAT USE ............................................................................................... 10 MAPPING EUROPEAN BISON HABITAT IN GERMANY ............................................................................................ 10 APPROACH ................................................................................................................................................................. -

FOREIGN RIGHTS Non Fiction London Preview 2020 Knesebeck Verlag | Holzstrasse 26 | 80469 Muenchen | Germany
FOREIGN RIGHTS non fiction London preview 2020 Knesebeck Verlag | Holzstrasse 26 | 80469 Muenchen | Germany T +49 (0) 89 242 11 66-0 | [email protected] | www.knesebeck-verlag.de The Fascination of Research "AT THE END OF A SUCCESSFUL DAY, YOU MIGHT HAVE CHANGED THE WORLD BECAUSE YOU DISCOVERED NEW KNOWLEDGE AND MADE IT ACCESSIBLE TO MANKIND." – DAVID AVNIR (CHEMIKER) For her latest project, well-known photographer Herlinde Koelbl visited top scientists worldwide, who outlined their areas of research and related their experiences. Each researcher sketched the basics of his or her work on the palm of their hand, making science tangible in the truest sense of the word! A photographic project which conveys the fascination of science in an unusually accessible way and offers a unique insight into the world of science, where men and women, independent of their origin and nationality, are driven by the same motivations: curiosity and the thirst for knowledge. THE AUTHOR Herlinde Koelbl studied fashion in Munich and only became a photographer in 1976. She has worked for well- draft cover known newspapers and magazines such as Stern, Die Zeit Photographer/Author: Herlinde Koelbl and New York Times. Her first broad success came with the publication of the photo book Das deutsche Wohnzimmer (The German Living Room) (1980), which was followed by other works. From 1991 to 1998, for her biggest project to date, a long-term study, she photographed and interviewed fifteen leading figures from the worlds of politics and business once a year, producing the photographic volume Spuren der Macht (Traces of Power) and an exhibition of the same name. -

Indicators of Hemeroby for the Monitoring of Landscapes in Germany
Indicators to monitor the structural diversity of landscapes Ulrich Walz Leibniz Institute of Ecological Urban and Regional Development, Weberplatz 1, 01217 Dresden, Germany Ecological Modelling 295 (2015) 88–106, http://dx.doi.org/10.1016/j.ecolmodel.2014.07.011 ABSTRACT An important level of biodiversity, alongside the diversity of genes and species, is the diversity of ecosystems and landscapes. In this contribution an indicator system is proposed to measure natural diversity (relief, soils, waters), cultural diversity (main land use classes, diversity of land use, ecotones, connectivity) and anthropogenic impacts (fragmentation, hemeroby, protection).The contribution gives an overview of various indicators on landscape diversity and heterogeneity currently used in Germany andEurope. Based on these indicators a complementary system, is presented. The indicators introduced here are derived from regular evaluations of the digital basis landscape model (BasicDLM) of the Authoritative Topographic-Cartographic Information System (ATKIS), the digital land cover model for Germany (LBM-DE) as well as other supplementary data such as the mapping of potential natural vegetation. With the proposed indicators it is possible to estimate cumulative land-use change and its impact on the environmental status and biodiversity, so that existing indicator systems are supplemented with meaningful additional information. Investigations have shown that indicators on forest fragmentation, hemeroby or ecotones can be derived from official geodata. As such geodata is regularly updated, trends in indicator values can be quickly identified. Large regional differences in the distribution of the proposed indicators have been confirmed, thereby revealing deficits and identifying those regions with a high potential for biodiversity. The indicators will be successively integrated into the web-based land-use monitor (http://www.ioer-monitor.de), which is freely available for public use. -

Biodiversity of the Hercynian Mountains of Central Europe
Pirineos, 151-152: 83 a 99, JACA; 1998 BIODIVERSITY OF THE HERCYNIAN MOUNTAINS OF CENTRAL EUROPE JAN JENÍK Faculty of Science, Charles University, Benátská 2, CZ-12801 Praha 2, Czech Republic SUMMARY.- The vegetation of temperate Central Europe north of the Alps is mainly of low diversity broadleaf and conifer forest. The occurrence of three azonal habitat types: mires with their numerous microhabitats, the georelief of the karst and its deeply-cut river valleys, and ecological islands with a distinct vegetation near the treeline of the middle-mountains causes local areas of high diversity. These high species diversity spots are the result of an interplay between physical, biotic and historical factors. A model of an anemo-orographic system with its underlying factors is described to explain the high plant and animal diversity in the corries (glacial cirques) of the Hercynian mountains. RÉSUMÉ.-La végétation de l'Europe Centrale tempérée au nord des Alpes nous montre surtout des forêts à de feuillues et de connifères à faible diversité. Cependant, la présence de trois types d'habitats azonaux peut produire une haute diversité au niveau local: zones humides avec leur nombreux microhabitats, reliefs karstiques et leur profonds défilés fluviaux, et enfin des îles écologiques avec une végétation par ticulière situées près de la limite supérieure des arbres (treeline). Ces secteurs riches en espèces peuvent s'expliquer par l'interaction des facteurs physiques, biotiques et historiques. Dans ce domaine nous proposons un système anémo-orographique avec ses facteurs inféodés qui pourrait expliquer la haute diversité animale et végétale dans les cirques glaciaires des montagnes hercyniennes. -

Proxy Environmental Record from the Bohemian Forest, Czech Republic
bs_bs_banner The Lateglacial and Holocene in Central Europe: a multi-proxy environmental record from the Bohemian Forest, Czech Republic KLARA VOCADLOV A, LIBOR PETR, PAVLA ZA CKOV A, MAREK KRIZEK, LENKA KRIZOV A, SIMON M. HUTCHINSON AND MIROSLAV SOBR Vocadlova, K., Petr, L., Zackova, P., Krızek, M., Krızova, L., Hutchinson, S. M. & Sobr M.: The Lateglacial and Holocene in Central Europe: a multi-proxy environmental record from the Bohemian Forest, Czech Republic. Boreas. 10.1111/bor.12126. ISSN 0300-9483. The Hercynian mountain ranges were islands of mountain glaciation and alpine tundra in a Central European ice-free corridor during the Late Pleistocene. Today they are notable areas of glacial landforms, alpine-forest free areas, peatlands and woodlands. However, our knowledge of the Lateglacial and early Holocene environmental changes in this region is limited. We present a new multi-proxy reconstruction of a mid-altitude environment in the Bohemian Forest spanning this period. A core (5.2 m length) in the Cern e Lake cirque (1028 m a.s.l.) was sub- jected to lithological, geochemical, pollen and macrofossil analysis supplemented by two optically stimulated luminescence (OSL) and 10 AMS radiocarbon dates. We determined the impact of regional and supraregional cli- mate changes on the environment. The two most significant changes in sedimentation during the Lateglacial (17.6 and 15.8–15.5 cal. ka BP) were synchronous with regional glacial chronostratigraphy. Unlike Central European mountain ranges, in the Bohemian Forest the Younger Dryas was not coincident with glacier re-advance, but was a dry, cold episode with low lake levels, which prevailed until the early Preboreal. -

Naturräumlich-Ökologische Analyse Der Flechtenflora Von Deutschland
624 Herzogia 28 (2) Teil 2, 2015: 624 – 653 Naturräumlich-ökologische Analyse der Flechtenflora von Deutschland Ulf Schiefelbein*, Florian Jansen, Birgit Litterski & Volkmar Wirth Zusammenfassung: Schiefelbein, U., Jansen, F., Litterski, B. & Wirth, V. 2015. Naturräumlich-ökologische Analyse der Flechtenflora von Deutschland. – Herzogia 28: 624 – 653. Die Flechtenflora von Deutschland wird auf der Grundlage der Angaben von Wirth et al. (2013; Die Flechten Deutschlands) analysiert, wobei Naturräume die geografische Basis für die Analysen bilden. Bewertet werden Artendiversität, Exklusivität des Arteninventars, substratspezifische Eigenschaften (Substratbindung, pH-Werte und Nährstoffgehalt/Eutrophierung der besiedelten Substrate) und klimatische Faktoren (Licht, Luftfeuchte). Die artenreichs- ten Naturräume sind nach den Bayerischen Alpen, dem Schwarzwald und Odenwald-Spessart die ebenfalls sehr nieder- schlagsreichen Naturräume Eifel, Weserbergland, Harz, Fränkische Alb, Sauerland und Bayerisch-Böhmischer Wald. Die artenärmsten Landschaften liegen überwiegend im südlichen Teil des Nordostdeutschen Tieflandes. Die Exklusivität des Arteninventars eines Naturraumes wird als Anzahl der Arten, die in Deutschland nach 1950 nur in einem bis zwei Naturräumen nachgewiesen wurden, definiert. In der gesamten Bundesrepublik sind es 638 Arten, davon kommen die meisten in den Bayerischen Alpen, im Schwarzwald, Bayerischen Wald, Odenwald-Spessart und in der Schwäbischen Alb vor. Im gesamten Deutschland überwiegen die Gesteinsbewohner (47,6 % des Gesamtarteninventars), -

2.14 Mean Annual Climatic Water Balance
2.14 Mean Annual Climatic Water Balance The climatic water balance (CWB) is defined as the difference between precipitation depth Baltic Sea. The whole lowland regions of Mecklenburg-Vorpommern (Mecklenburg-Western and the depth of potential evapotranspiration at a given site during a certain time period. Pomerania), Brandenburg, Sachsen-Anhalt (Saxony-Anhalt), and Sachsen (Saxony) have negative summer half-year balances, with average values sometimes drastically below In general climatology, climate classifications are usually based on the weather elements “air - 100 mm. The highest deficits in the summer half-year show values below -300 mm. In sum- temperature” and “precipitation depth”, from which e. g. the description of the aridity of the mers with abundant rainfall, positive half-year balances may be recorded too, what was the climate is derived, the so-called aridity index. However, in the context of water-resources case in about one third of the years in the series 1961–1990. management and hydrology, the climatic water balance is better suitable for the hydroclimatic characterisation of sites, areas or periods, because the (hydro-)climatic conditions are The period with mean negative monthly balances in the inland lowlands lasts from April to described directly by means of the water-balance effective elements “precipitation” or “poten- September/October. The highest monthly balance deficits below -100 mm are recorded in the tial evapotranspiration” in the dimension “mm”. Dependent on whether precipitation depth or months from May to July. Negative monthly balances may occur throughout the year, potential evapotranspiration depth prevails in the considered period, the climatic water provided dry weather prevails. -

Sulphate Sorption Characteristics of the Bohemian Forest Soils
Silva Gabreta vol. 11 (1) p. 3–12 Vimperk, 2005 Sulphate sorption characteristics of the Bohemian Forest soils Jiří Kaňa* & Jiří Kopáček Faculty of Biological Sciences, University of South Bohemia, and Hydrobiological Institute, AS CR, Na Sádkách 7, CZ-37005 České Budějovice, Czech Republic *[email protected] Abstract Sulphate (SO4-S) sorption characteristics were determined in soils from catchments of three acidified mountain lakes, Plešné (PL), Čertovo (CT), and Černé (CN), situated in the Bohemian Forest. The SO4-S desorption ranged between 0.07–0.73, 0.21–0.68, and 0.39–0.82 mmol.kg–1 in the PL, CT, and CN catchments, respectively. SO4-S desorption was observed to be higher in horizon A than in mineral horizons below. The SO4-S represented only 3.4% of total S pool in the catchments and did not explain a relatively high rate of terrestrial leaching of sulphate over last 10 years. This disproportion suggests that the soil SO4-S pool is being replenished from internal soil processes from the pool of total S. Sulphate sorption maxima in soils in the catchments of the Bohemian Forest lakes varied between 1.5–11.7 mmol.kg–1, being highest in A-horizon and lowest in E-horizon. Key words: acidification, recovery, soil chemistry INTRODUCTION The chemistry (and, consequently, biology) of mountain lakes is determined by amount of precipitation and their composition, bedrock (PSENNER & CATALAN 1994), catchment soils and vegetation (e.g. BARON et al. 1994), and hydrology (WETZEL 2002). Soil sorption charac- teristics may significantly affect water composition, e.g. -

Vegetation of the Czech Republic: Diversity, Ecology, History and Dynamics
Preslia 84: 427–504, 2012 427 Vegetation of the Czech Republic: diversity, ecology, history and dynamics Vegetace České republiky: diverzita, ekologie, historie a dynamika Dedicated to the centenary of the Czech Botanical Society (1912–2012) Milan C h y t r ý Department of Botany and Zoology, Masaryk University, Kotlářská 2, CZ-611 37 Brno, Czech Republic, e-mail: [email protected] Chytrý M. (2012): Vegetation of the Czech Republic: diversity, ecology, history and dynamics. – Preslia 84: 427–504. This review summarizes basic information on the diversity of vegetation in the Czech Republic. It describes basic environmenal factors affecting vegetation, vegetation history since the last glacial, biomes occurring in the Czech Republic (zonal biomes of broad-leaved deciduous forest and forest- steppe, and azonal biomes of taiga and tundra), altitudinal zonation of vegetation and landscapes with an exceptionally high diversity of vegetation types (deep river valleys in the Bohemian Massif, karst areas, sandstone pseudokarst areas, solitary volcanic hills, glacial cirques, lowland riverine landscapes and serpentine areas). Vegetation types, delimited according to the monograph Vegeta- tion of the Czech Republic, are described with emphasis on their diversity, ecology, history and dynamics. K e y w o r d s: alpine, aquatic, central Europe, forest, grassland, phytosociology, plant communi- ties, ruderal, vegetation change, vegetation classification, vegetation history, weed, wetland Introduction The Czech Republic is a land-locked country in central Europe occupying an area of 78,867 km2. It is situated in the zone of temperate broad-leaved deciduous forest, which in the south-east borders on the forest-steppe zone. -

Late Cretaceous to Paleogene Exhumation in Central Europe – Localized Inversion Vs
https://doi.org/10.5194/se-2020-183 Preprint. Discussion started: 11 November 2020 c Author(s) 2020. CC BY 4.0 License. Late Cretaceous to Paleogene exhumation in Central Europe – localized inversion vs. large-scale domal uplift Hilmar von Eynatten1, Jonas Kley2, István Dunkl1, Veit-Enno Hoffmann1, Annemarie Simon1 1University of Göttingen, Geoscience Center, Department of Sedimentology and Environmental Geology, 5 Goldschmidtstrasse 3, 37077 Göttingen, Germany 2University of Göttingen, Geoscience Center, Department of Structural Geology and Geodynamics, Goldschmidtstrasse 3, 37077 Göttingen, Germany Correspondence to: Hilmar von Eynatten ([email protected]) Abstract. Large parts of Central Europe have experienced exhumation in Late Cretaceous to Paleogene time. Previous 10 studies mainly focused on thrusted basement uplifts to unravel magnitude, processes and timing of exhumation. This study provides, for the first time, a comprehensive thermochronological dataset from mostly Permo-Triassic strata exposed adjacent to and between the basement uplifts in central Germany, comprising an area of at least some 250-300 km across. Results of apatite fission track and (U-Th)/He analyses on >100 new samples reveal that (i) km-scale exhumation affected the entire region, (ii) thrusting of basement blocks like the Harz Mountains and the Thuringian Forest focused in the Late 15 Cretaceous (about 90-70 Ma) while superimposed domal uplift of central Germany is slightly younger (about 75-55 Ma), and (iii) large parts of the domal uplift experienced removal of 3 to 4 km of Mesozoic strata. Using spatial extent, magnitude and timing as constraints suggests that thrusting and crustal thickening alone can account for no more than half of the domal uplift. -

Eastern Bavaria
Basic text Eastern Bavaria Culture Eastern Bavaria is still home to more castles than anywhere else in Germany: Some medieval castles remain only as ruins, whilst other castles such as Falkenstein Castle have withstood decline and are open to visitors. The expansive spruce forests in Eastern Bavaria have given way to the Bavarian Glass Road, as they supplied the wood and quartz sand –the key raw materials – for the very first glass foundries. Spanning some 250 kilometres, it is one of the most picturesque holiday routes in Germany. Those choosing to travel along the route will learn all about the 700-year tradition of glass production and glass as a form of art. The route, which begins in Neustadt an der Waldnaab and leads to Passau, features glass foundries, galleries and museums, all packed to the brim with interesting facts about the traditional handicraft. Some Eastern Bavarian companies are keeping the tradition alive to this day and export to countries ranging from the United Arab Emirates to the United States of America. The largest towns in Eastern Bavaria include Regensburg, Landshut and Passau. The city of Regensburg, which was first founded by Roman Emperor Marcus Aurelius, has retained its medieval centre to this day. The Old Town of Regensburg together with Stadtamhof has been a UNESCO World Heritage Site since 2006. Landshut is the prototype of an old Bavarian town. Above all its town centre, which features gabled houses, decorative façades, oriels and arches, is one of the most beautiful squares to be found in the whole of Germany. The three-river town of Passau, which was built in the Italian baroque style, achieved early wealth thanks to its participation in the salt trade and was a place of border crossings due to its location on the border with Austria and just 30 kilometres from the Czech border. -
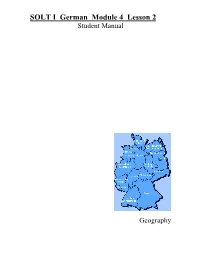
SOLT I German Module 4 Lesson 2 Student Manual
SOLT I German Module 4 Lesson 2 Student Manual Geography Geography German SOLT I Objectives Module 4 Lesson 2 At the end of this lesson you will be able identify the different German states and the countries surrounding Germany. In order to achieve this objective you will: Recognize the German States and Their Locations • Recognize the states’ locations, their capitals, major cities, and regions within the states • Describe their areas and borders • Brief about Germany • Describe the geography of Germany • Talk about the location of military installations in Germany • Identify the national flag of the country Identify Nationalities • Identify languages in the world countries • Identify different ethnic groups • Describe demographic composition 47 Geography German SOLT I Introduction Module 4 Lesson 2 Features of Germany You are planning a short vacation somewhere in Germany and have asked one of your German counterparts to describe some of the various landscapes, since you can’t decide. He gives you a quick description of Germany’s geography. Where will you decide to go? Deutschland hat sechzehn Bundesländer. Die Hauptstadt von Deutschland ist Berlin. In jedem Bundesland gibt es sehr schöne Landschaften. Im Norden ist die Nordsee mit der schönen Insel Sylt. Das Land ist flach. Die Bundesländer dort sind Schleswig-Holstein, Hamburg und Bremen. In der Mitte von Deutschland gibt es viele Berge. Besonders schön ist der Teutoburger Wald und der Hunsrück. Weiter südlich gibt es natürlich den berühmten Schwarzwald im Westen und den Bayrischen Wald. Einige Bundesländer dort sind Sachsen-Anhalt, Thüringen, Hessen und Nordrhein-Westfalen. Im südlichen Teil von Deutschland liegt Bayern.