Shrub Encroachment of Temperate Grasslands: Effects on Plant
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

University of Cape Town
The copyright of this thesis vests in the author. No quotation from it or information derived from it is to be published without full acknowledgementTown of the source. The thesis is to be used for private study or non- commercial research purposes only. Cape Published by the University ofof Cape Town (UCT) in terms of the non-exclusive license granted to UCT by the author. University Some consequences of woody plant encroachment in a mesic South African savanna Emma Fiona Gray Town Cape of Submitted in fulfilment of the requirements for a Master of Science Degree Supervisor UniversityProfessor William Bond Botany Department University of Cape Town Rondebosch 7701 March 2011 Acknowledgements My most sincere thanks go to my supervisor, Professor William Bond. There is no doubt that without his guidance, belief and support over the last two years I would still be sitting in front of a blank screen. Thanks for the passion, the inspiration and all the amazing opportunities. Thanks to Ezemvelo KZN Wildlife for allowing me to conduct my research in their park. Particular thanks go to Dr Dave Druce, for welcoming me into his research centre and giving me the staff and freedom I needed during my fieldwork. I am also extremely grateful to Geoff Clinning for his patience and willingness to help with all my data and GIS needs. Thanks to the Zululand Tree Project for logistical support during my field work. To my field assistants, Njabulo, Bheki and Ncobile, for their uncomplaining hard work in the most trying conditions. Without them I would have no data. -

Fire Dynamics in Mallee-Heath Fuel, Weather and Fire Behaviour Prediction in South Australian Semi-Arid Shrublands
PROGRAM A REPORT NO. A.10.01 FIRE DYNAMICS IN MALLEE-HEATH FUEL, WEATHER AND FIRE BEHAVIOUR PREDICTION IN SOUTH AUSTRALIAN SEMI-ARID SHRUBLANDS M.G. Cruz1, S. Matthews1, J. Gould1, P. Ellis1, M. Henderson2, I. Knight1, J. Watters1 1 Bushfire Dynamics and Applications, CSIRO Sustainable Ecosystems and CSIRO Climate Adaptation Flagship, Canberra, ACT, Australia 2 Department of Environment and Heritage, Adelaide, SA, Australia © BUSHFIRE CRC LTD 2006 © Bushfire Cooperative Research Centre 2006. No part of this publication must be reproduced, stored in a retrieval system or transmitted in any form without prior written permission from the copyright owner, except under the conditions permitted under the Australian Copyright Act 1968 and subsequent amendments. Publisher: CSIRO Sustainable Ecosystems Canberra, ACT, Australia ISBN: 0 00000 000 0 March 2010 PROGRAM A REPORT NO. A.10.01 FIRE DYNAMICS IN MALLEE-HEATH FUEL, WEATHER AND FIRE BEHAVIOUR PREDICTION IN SOUTH AUSTRALIAN SEMI-ARID SHRUBLANDS M.G. Cruz1, S. Matthews1, J. Gould1, P. Ellis1, M. Henderson2, I. Knight1, J. Watters1 1 Bushfire Dynamics and Applications, CSIRO Sustainable Ecosystems and CSIRO Climate Adaptation Flagship, Canberra, ACT, Australia 2 Department of Environment and Heritage, Adelaide, SA, Australia © BUSHFIRE CRC LTD 2006 PROGRAM A :: REPORT NO. A.10.01 CONTENTS ABSTRACT - FIRE DYNAMICS IN MALLEE-HEATH 10H ACKNOWLEDGEMENTS 21H EXECUTIVE SUMMARY 32H 2. EXPERIMENTAL DESIGN 63H 2.1. Site characteristics 64H 2.2. Fire climate 95H 2.3. Plot layout 136H 2.4. Methods 147H 3. FUEL COMPLEX DYNAMICS 198H 3.1. Introduction 199H 3.2. Methods 2210H 3.3. Results 241H 3.4. Discussion and conclusions 3412H 4. -

3-2-Effects-Of-Fire-Regime-On-Plant
Foster, C. N., Barton, P. S., MacGregor, C. I., Catford, J. A., Blanchard, W., & Lindenmayer, D. B. Effects of fire regime on plant species richness and composition differ among forest, woodland and heath vegetation. Applied Vegetation Science, 21(1): 132-143. DOI: https://doi.org/10.1111/avsc.12345 Page 1 of 29 Applied Vegetation Science EFFECTS OF FIRE REGIME ON PLANT SPECIES RICHNESS AND COMPOSITION DIFFER AMONG FOREST, WOODLAND AND HEATH VEGETATION Foster, C.N. (corresponding author, [email protected])1,2 Barton, P.S. ([email protected])1 MacGregor, C.I. ([email protected])1,2,3 Catford, J.A. ([email protected])1,2,4,5 Blanchard, W. ([email protected]) 1 Lindenmayer, D.B. ([email protected]) 1,2,3 1 Fenner School of Environment and Society, The Australian National University, Canberra, ACT, 2601, Australia 2Australian Research Council Centre of Excellence for Environmental Decisions, The Australian National University, Canberra, ACT, 2601, Australia 3The National Environmental Science Program, Threatened Species Recovery Hub and the Long-term Ecological Research Network, Fenner School of Environment and Society, The Australian National University, Canberra, ACT, 2601, Australia 4School of BioSciences, The University of Melbourne, Parkville, Vic, 3010, Australia 5Biological Sciences, University of Southampton, Highfield Campus, Southampton, SO17 1BJ, UK. Keywords: community composition, competition, disturbance regime, dry sclerophyll vegetation, fire management, fire frequency, Sydney Coastal Heath, Sydney Coastal Forest, species richness Nomenclature: Harden (1991) for species, Taws (1997) for plant communities Running Head: Fire regimes in dry sclerophyll vegetation Applied Vegetation Science Page 2 of 29 1 ABSTRACT 2 Question: Do the effects of fire regimes on plant species richness and composition differ among 3 floristically similar vegetation types? 4 Location: Booderee National Park, south-eastern Australia. -

Natura 2000 Interpretation Manual of European Union
NATURA 2000 INTERPRETATION MANUAL OF EUROPEAN UNION HABITATS Version EUR 15 Q) .c Ol c: 0 "iii 0 ·"'a <>c: ~ u.. C: ~"' @ *** EUROPEAN COMMISSION ** ** DGXI ... * * Environment, Nuclear Security and Civil Protection 0 *** < < J J ) NATURA 2000 INTERPRETATION MANUAL OF EUROPEAN UNION HABITATS Version EUR 15 This 111anual is a scientific reference document adopted by the habitats committee on 25 April 1996 Compiled by : Carlos Romio (DG. XI • 0.2) This document is edited by Directorate General XI "Environment, Nuclear Safety and Civil Protection" of the European Commission; author service: Unit XI.D.2 "Nature Protection, Coastal Zones and Tourism". 200 rue de Ia Loi, B-1049 Bruxelles, with the assistance of Ecosphere- 3, bis rue des Remises, F-94100 Saint-Maur-des-Fosses. Neither the European Commission, nor any person acting on its behalf, is responsible for the use which may be made of this document. Contents WHY THIS MANUAL?---------------- 1 Historical review ............................................... 1 The Manual .................................................... 1 THE EUR15 VERSION 3 Biogeographical regions .......................................... 3 Vegetation levels ................................................ 4 Explanatory notes ............................................... 5 COASTAL AND HALOPHYTIC HABITATS 6 Open sea and tidal areas . 6 Sea cliffs and shingle or stony beaches ............................ 10 Atlantic and continental salt marshes and salt meadows . 12 Mediterranean and thermo-Atlantic saltmarshes and salt meadows .... 14 Salt and gypsum continental steppes . 15 COASTAL SAND DUNES AND CONTINENTAL DUNES 17 Sea dunes of the Atlantic, North Sea and Baltic coasts ............... 17 Sea dunes of the Mediterranean coast . 22 Continental dunes, old and decalcified . 24 FRESHWATER HABITATS 26 Standing water . 26 Running water . 29 TEMPERATE HEATH AND SCRUB------------ 33 SCLEROPHYLLOUS SCRUB (MATORRAL) 40 Sub-Mediterranean and temperate . -

Ecological Distribution of Four Co-Occurring Mediterranean Heath Species
ECOGRAPHY 23: 148–159. Copenhagen 2000 Ecological distribution of four co-occurring Mediterranean heath species Fernando Ojeda, Juan Arroyo and Teodoro Maran˜o´n Ojeda, F., Arroyo, J. and Maran˜o´n, T. 2000. Ecological distribution of four co-occurring Mediterranean heath species. – Ecography 23: 148–159. Erica australis, E. scoparia, E. arborea and Calluna ulgaris are the most abundant heath species on acid, sandstone-derived soils of the Strait of Gibraltar region (southern Spain and northern Morocco). Despite their apparently similar ecological requirements, these four species are somewhat ecologically segregated. Erica australis is abundant only on poor, shallow soils, with a high content in soluble aluminium, generally on mountain ridges and summits. Erica scoparia becomes dominant on deeper sandstone soils with lower aluminium. Calluna ulgaris coexists with these Erica species in communities under low or no tree cover. In the Spanish side of the Strait (Algeciras), Erica arborea tends to be relegated to communities under moderate to dense tree cover, whereas this species is more abundant and widespread in the Moroccan side (Tangier). Tolerance to extreme physical conditions – high aluminium and dense tree cover – and interspecific competition seem to explain the ecological distribution of these four heath species in the Strait of Gibraltar region. The more fragmented pattern of sandstone patches and higher disturbance levels in Tangier might account for the differences in the patterns of ecological distribution of these four heath species between both sides of the Strait of Gibraltar. F. Ojeda ([email protected]) and J. Arroyo, Dept de Biologı´a Vegetal y Ecologı´a, Uni. de Seilla. -
Vegetation Formations, Classes and Communities
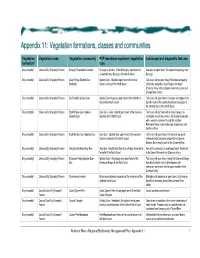
Appendix 11: Vegetatiion formatiions, cllasses and communiities Vegetation Vegetation class Vegetation community PVP developer equivalent vegetation Landscape and diagnostic features formation* type Dry sclerophyll Clarence Dry Sclerophyll Forests Baryulgil Serpentinite Complex Eucalyptus ophitica - White Mahogany open forest on Low open or open forest. On serpentinite geology near serpentinite near Baryulgil of the North Coast Baryulgil. Dry sclerophyll Clarence Dry Sclerophyll Forests Coast Range Spotted Gum- Spotted Gum - Blackbutt open forest of the lower Tall to very tall dry open forest. Restricted and patchy Blackbutt Clarence Valley of the North Coast distribution along the Coast Range in the lower Clarence Valley, with a disjunct western occurrence in Grange State Forest. Dry sclerophyll Clarence Dry Sclerophyll Forests Dry Foothills Spotted Gum Spotted Gum dry grassy open forest of the foothills of Tall to very tall open forest. On slopes and ridges of dry the northern North Coast foothills areas of the coastal hinterland and gorges of the northern parts of the North Coast. Dry sclerophyll Clarence Dry Sclerophyll Forests Foothill Grey Gum-Ironbark- Grey Gum - Grey Ironbark open forest of the Clarence Tall to very tall dry forest with a mixed canopy. On Spotted Gum lowlands of the North Coast sandstone and siliceous soils in the Clarence lowlands with a western extension through the southern Richmond Range inland to Ewingar State Forest and the Mann River. Dry sclerophyll Clarence Dry Sclerophyll Forests Foothills Grey Gum-Spotted Gum Grey Gum - Spotted Gum open forest of the southern Tall to very tall open forest. On high and low quartz Clarence lowlands of the North Coast sediments in the southern portion of the Clarence- Moreton Basin mainly south of the Clarence River. -

Habitat Islands in Fire-Prone Vegetation: Do Landscape Features
Journal of Biogeography, 29, 677–684 Habitat islands in fire-prone vegetation: do landscape features influence community composition? Peter J. Clarke Department of Botany, University of New England, Armidale, Australia Abstract Aim, Location Landscape features, such as rock outcrops and ravines, can act as habitat islands in fire-prone vegetation by influencing the fire regime. In coastal and sub- coastal areas of Australia, rock outcrops and pavements form potential habitat islands in a matrix of fire-prone eucalypt forests. The aim of this study was to compare floristic composition and fire response traits of plants occurring on rocky areas and contrast them with the surrounding matrix. Methods Patterns of plant community composition and fire response were compared between rocky areas and surrounding sclerophyll forests in a range of climate types to test for differences. Classification and ordination were used to compare floristic composition and univariate analyses were used to compare fire response traits. Results The rock outcrops and pavements were dissimilar in species composition from the forest matrix but shared genera and families with the matrix. Outcrops and pavements were dominated by scleromorphic shrubs that were mainly killed by fire and had post-fire seedling recruitment (obligate seeders). In contrast, the most abundant species in the adjacent forest matrix were species that sprout after fire (sprouters). Main conclusions Fire frequency and intensity are likely to be less on outcrops than in the forest matrix because the physical barrier of rock edges disrupts fires. Under the regime of more frequent fires, obligate seeders have been removed or reduced in abundance from the forest matrix. -

Guidelines for Ecologically Sustainable Fire
GGUUIIDDEELLIINNEESS FFOORR EECCOOLLOOGGIICCAALLLLYY SSUUSSTTAAIINNAABBLLEE FFIIRREE MMAANNAAGGEEMMEENNTT NSW BIODIVERSITY STRATEGY JULY 2004 GGUUIIDDEELLIINNEESS FFOORR EECCOOLLOOGGIICCAALLLLYY SSUUSSTTAAIINNAABBLLEE FFIIRREE MMAANNAAGGEEMMEENNTT A project undertaken for the NSW Biodiversity Strategy For more information and for access to the databases contact: Bushfire Research Unit, Biodiversity Research & Management Division NSW National Parks and Wildlife Service,PO Box 1967, Hurstville, NSW 2220 Ph. (02) 9585 6643, Fax (02) 9585 6606 Website: www.npws.nsw.gov.au © Crown copyright March 2003 New South Wales Government ISBN 0731367022 This project has been funded by the NSW Biodiversity Strategy and the NSW National Parks & Wildlife Service and carried out by the following staff from the Biodiversity Research and Management Division (Bushfire Research), NPWS: Belinda Kenny Elizabeth Sutherland Elizabeth Tasker Ross Bradstock Photograph by Elizabeth Tasker Disclaimer While every reasonable effort has been made to ensure that this document is correct at the time of printing, the State of New South Wales, its agents and employees, do not assume any responsibility and shall have no liability, consequential or otherwise, of any kind, arising from the use of or reliance on any of the information contained in this document. CONTENTS Project summary 1: Introduction 9 1.1: Background 9 1.2: Limitations 14 2: Methodology 16 2.1: Overview of approach 16 2.2: Fire response databases 17 2.3: Summarising the fire response databases 21 2.4: Allocation -
Mediterranean Biome
Mediterranean Biome Mediterranean Biome § 30° to 40° N and S latitude on the west sides of continents “ § Just poleward of the subtropical deserts on the western The similarity of form and functional continental edges. response of the vegetation to the rigorous mediterranean environment is therefore a striking example of evolutionary convergence, and has resulted in a high degree of endemism within the regional floras" (Archibold, 1995) Mediterranean Biome Mediterranean Biome § subtropical dry and warm air in summer, cold currents § the Mediterranean biome is sandwiched § in winter, as subtropical highs retreat toward equator, they between deserts and temperate rainforests experience maritime airmasses and cyclonic storms from polar on west sides of continents - experience front both but in alternating seasons Biome types on west side of Chile Desert Mediterranean Mediterranean Desert Temperate rainforest 'winter-rain and summer dry' 1 Mediterranean Biome Mediterranean Floristic Regions § Californian § Capensic (South African) Vancouver - approaching temperate rainforest § Mediterranean § Australian (slight summer dry § Chilean period) Pasadena - classic Mediterranean climate (6 months rain, 6 months dry) San Diego - shift to more desert conditions (reduced winter rain) Mediterranean Floristic Regions Mediterranean Vegetation § Californian § Capensic (South African) § the Mediterranean Biome and its vegetation is closely linked with fire ecology § Mediterranean § Australian § Chilean What do these sound like? Wines! Santa Barbara chapparal -
Fuel and Fire Behaviour in Semi-Arid Mallee-Heath Shrublands
VI International Conference on Forest Fire Research D. X. Viegas (Ed.), 2010 Fuel and fire behaviour in semi-arid mallee-heath shrublands M.G. Cruz, J.S. Gould 1 CSIRO Ecosystem Sciences and CSIRO Climate Adaptation Flagship - Bushfire Dynamics and Applications, Canberra ACT Australia ([email protected]) 2 Bushfire Cooperative Research Centre, East Melbourne VIC Australia Abstract An experimental burning program was set up in South Australia aimed at characterizing fuel dynamics and fire behaviour in mallee-heath woodlands. Fuel complexes in the experimental area comprised mallee and heath vegetation with ages (time since fire) ranging from 7 to 50 years old. Dominant overstorey mallee vegetation comprised Eucalyptus calycogona , E. diversifolia , E. incrassate and E. leptophylla . A total of 66 fires were completed. The range of fire environment conditions within the experimental fire dataset were: air temperature 15 to 39°C; relative humidity 7 to 80%; mean 10-m open wind speed 3.6 to 31.5 km/h; Forest Fire Danger Index 1.7 to 46. Total fuel load ranged from 0.38 kg/m 2 in young (7-year old) mallee to 1.0 kg/m 2 in mature stands. Fire behaviour measurements included rate of spread, flame geometry, residence time and fuel consumption. Measured rate of spread ranged between 0.8 and 55 m/min with fireline intensity between 144 and 11,000 kW/m. The dataset provided insight into the threshold environment conditions necessary for the development of a coherent flame front able to overcome the fine scale fuel discontinuities that characterise the semi-arid mallee-heath fuel types and support self-sustained fire propagation. -

Tasmania East Coast Conservation Corridor
East Coast Conservation Corridor A landscape of national significance North East Bioregional Network October 2012 Report produced for the North East Bioregional Network www.northeastbioregionalnetwork.org.au October 2012 Nick Fitzgerald BSc (Hons) [email protected] All photographs by the author 1 Contents Summary.......................................................................................................................................................................... 3 Introduction.................................................................................................................................................................... 6 Geology and Geomorphology............................................................................................................................. 7 Climate........................................................................................................................................................................ 7 Climate Change........................................................................................................................................................ 8 Land Tenure.............................................................................................................................................................. 9 Reserves................................................................................................................................................................ 9 State Forest and Crown Land.................................................................................................................... -

A Hierarchical Classification of Wildland Fire Fuels for Australian
fire Concept Paper A Hierarchical Classification of Wildland Fire Fuels for Australian Vegetation Types Miguel G. Cruz 1,* ID , Jim S. Gould 1, Jen J. Hollis 1,2 and W. Lachie McCaw 3 1 CSIRO, GPO Box 1700, Canberra, ACT 2601, Australia; [email protected] (J.S.G.); [email protected] (J.J.H.) 2 New South Wales Rural Fire Service, P.O. Box 2234, Queanbeyan, NSW 2620, Australia 3 Department of Parks and Wildlife, Manjimup, WA 6258, Australia; [email protected] * Correspondence: [email protected] Received: 19 March 2018; Accepted: 11 April 2018; Published: 17 April 2018 Abstract: Appropriate categorisation and description of living vegetation and dead biomass is necessary to support the rising complexity of managing wildland fire and healthy ecosystems. We propose a hierarchical, physiognomy-based classification of wildland fire fuels—the Bushfire Fuel Classification—aimed at integrating the large diversity of Australian vegetation into distinct fuel types that are easily communicated and quantitatively described. At its basis, the classification integrates life form characteristics, height, and foliage cover. The hierarchical framework, with three tiers, describes fuel types over a range of application requirements and fuel description accuracies. At the higher level, the fuel classification identifies a total of 32 top-tier fuel types divided into 9 native forest or woodland, 2 plantation, 10 shrubland, 7 grassland, and 4 other fuel types: wildland urban interface areas, horticultural crops, flammable wetlands, and nonburnable areas. At an intermediate level, the classification identifies 51 mid-tier fuel types. Each mid-tier fuel type can be divided into 4 bottom-tier fuel descriptions.