EFFECTS of COW GREEN RESERVOIR UPON DOWNSTREAM FISH POPULATIONS D. T. CRISP Introduction in 1967 the FBA Began Pre-Impoundment S
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
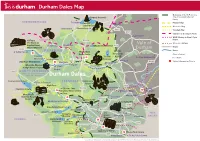
Durham Dales Map
Durham Dales Map Boundary of North Pennines A68 Area of Outstanding Natural Barleyhill Derwent Reservoir Newcastle Airport Beauty Shotley northumberland To Hexham Pennine Way Pow Hill BridgeConsett Country Park Weardale Way Blanchland Edmundbyers A692 Teesdale Way Castleside A691 Templetown C2C (Sea to Sea) Cycle Route Lanchester Muggleswick W2W (Walney to Wear) Cycle Killhope, C2C Cycle Route B6278 Route The North of Vale of Weardale Railway England Lead Allenheads Rookhope Waskerley Reservoir A68 Mining Museum Roads A689 HedleyhopeDurham Fell weardale Rivers To M6 Penrith The Durham North Nature Reserve Dales Centre Pennines Durham City Places of Interest Cowshill Weardale Way Tunstall AONB To A690 Durham City Place Names Wearhead Ireshopeburn Stanhope Reservoir Burnhope Reservoir Tow Law A690 Visitor Information Points Westgate Wolsingham Durham Weardale Museum Eastgate A689 Train S St. John’s Frosterley & High House Chapel Chapel Crook B6277 north pennines area of outstanding natural beauty Durham Dales Willington Fir Tree Langdon Beck Ettersgill Redford Cow Green Reservoir teesdale Hamsterley Forest in Teesdale Forest High Force A68 B6278 Hamsterley Cauldron Snout Gibson’s Cave BishopAuckland Teesdale Way NewbigginBowlees Visitor Centre Witton-le-Wear AucklandCastle Low Force Pennine Moor House Woodland ButterknowleWest Auckland Way National Nature Lynesack B6282 Reserve Eggleston Hall Evenwood Middleton-in-Teesdale Gardens Cockfield Fell Mickleton A688 W2W Cycle Route Grassholme Reservoir Raby Castle A68 Romaldkirk B6279 Grassholme Selset Reservoir Staindrop Ingleton tees Hannah’s The B6276 Hury Hury Reservoir Bowes Meadow Streatlam Headlam valley Cotherstone Museum cumbria North Balderhead Stainton RiverGainford Tees Lartington Stainmore Reservoir Blackton A67 Reservoir Barnard Castle Darlington A67 Egglestone Abbey Thorpe Farm Centre Bowes Castle A66 Greta Bridge To A1 Scotch Corner A688 Rokeby To Brough Contains Ordnance Survey Data © Crown copyright and database right 2015. -

Download Chapter In
Freshwater life Martyn G Kelly, Trevor D Cris, Ben Lamb and Brian Whitton Introduction Without the River Tees there would be no Teesdale and so, whilst much of the attention in this book is focussed on the plant and animal life in the fields and fells, we should not ignore either the river or the numerous tributary streams that feed it. Cow Green Reservoir, too, plays an important part in the story of Upper Teesdale, not just because of the ways in which it has altered the landscape and habitats in the upper valley, but also because the decision to impound the river precipitated many significant ecological studies and, ironically, raised the profile of the Teesdale rarities beyond a small band of botanical cognoscenti. The River Tees was the first British River to receive a detailed biological survey (by Butcher and colleagues in the mid-1930s). This was followed in the 1970s and 1980s by studies of the upper reaches of the main river and its tributaries by Durham University and the Freshwater Biological Association (later Institute of Freshwater Ecology and now Centre for Ecology and Hydrology). Since the previous edition of this book, further studies have investigated a wide range of factors including gravel, heavy metals, availability of salmonid spawning habitat and water colour. The upper tributaries of the Tees range from torrential streams, fed at times of peak flow mainly by surface run-off, to calcareous streams with some or much of their water from limestone springs. Those with the most water from springs are the ones which vary least in flow and have the highest calcium concentrations. -

Low Force and Holwick Are in the North Pennines Area of Outstanding Natural Beauty (AONB) and Geology and Landscape Around European Geopark
Low Force and Holwick are in the North Pennines Area of Outstanding Natural Beauty (AONB) and Geology and landscape around European Geopark European Geoparks The North Pennines AONB is Britain’s first European Low Force Geopark, a status supported by UNESCO, and a founding member of the Global Geoparks Network. Geoparks are special places with outstanding geology and landscape, and Holwick and where there are strong local efforts to make the most of geological heritage through interpretation, education, conservation and nature tourism. To find out more visit www.europeangeoparks.org A 2½-mile walk exploring landscape, Walk starts from here rocks, fossils and mines North Pennines Moor House – Upper Teesdale National Nature AONB & European Reserve (NNR) Geopark © Crown Copyright. All rights reserved. Part of this walk (south of the River Tees near Low Force) is Durham County Council. LA100049055. 2011. within the Moor House – Upper Teesdale NNR. This large reserve contains an almost complete range of upland For more information please contact: habitats typical of the North Pennines, from hay meadows North Pennines AONB Partnership, +44 (0)1388 528801 and juniper woods to limestone grassland and blanket bog. Weardale Business Centre, [email protected] It also includes the waterfalls of Cauldron Snout and High The Old Co-op Building, www.northpennines.org.uk 1 Martin Street, Stanhope, twitter.com/NorthPennAONB Force. For more information contact the Reserve Base on Bishop Auckland, County Durham facebook.com/NorthPenninesAONB 01833 622374. DL13 2UY Find out more about North Pennine geology This leaflet is one of a series of geological publications about the North Pennines. -

HADRIAN HUNDRED 25Th - 27Th MAY 2019
HADRIAN HUNDRED 25th - 27th MAY 2019 REGISTRATION – QUEEN ELIZABETH HIGH SCHOOL, HEXHAM NY 926 639 Welcome to Hexham once the haunt of marauding Vikings but now England’s favourite market town with the imposing Abbey at its hub. Starting in Northumberland the route visits Cumbria and Durham before returning to Northumberland for the later stages. Highlights include sections on Hadrian’s Wall, the South Tyne Trail, the Pennine Way (with Cross Fell and High Cup Nick), the Weardale Way and Isaac’s Tea Trail. Abbreviations TR Turn Right TL Turn Left N, S North, South etc. XXXm,Xkm Approx. distance in metres or kilometres to next feature (XXXdeg) Approx. magnetic bearing in degrees to next feature Units Convention Stage Summaries Miles & Kilometres (Distance), Feet & Metres (Ascent) Descriptive Text Metres (m) & Kilometres (km) NB. 100 metres = 109 yards 1 Kilometre = 0.62 miles Please note that all measurements of distance and ascent are produced from a GPS device which gives good estimates only therefore great accuracy cannot be guaranteed. 1 Important Notes A significant proportion of the Route uses or crosses roads, the vast majority of which are very minor and little used. The modern approach to Risk Assessment, however, requires that the risks involved in potentially mixing foot and vehicular traffic are pointed out whenever this happens. It is not proposed to mention this on every occasion that it occurs in the Route Description narrative. When a road is used or crossed the appropriate description will be highlighted. Additional warnings will be given whenever more major roads are encountered. Please be vigilant on roads especially later in the Event as you become increasingly tired and possibly less attentive. -

Moor House - Upper Teesdale B6278 Widdybank Farm, Langdon Beck, River Tees NNR Forest-In-Teesdale, B6277 Barnard Castle, Moor House – Cow Green Middleton- Co
To Alston For further information A686 about the Reserve contact: A689 The Senior Reserve Manager Moor House - Upper Teesdale B6278 Widdybank Farm, Langdon Beck, River Tees NNR Forest-in-Teesdale, B6277 Barnard Castle, Moor House – Cow Green Middleton- Co. Durham DL12 0HQ. Reservoir in-Teesdale To Penrith Tel 01833 622374 Upper Teesdale Appleby-in- National Nature Reserve Westmorland B6276 0 5km B6260 Brough To Barnard Castle B6259 A66 A685 c Crown copyright. All rights reserved. Kirkby Stephen Natural England 100046223 2009 How to get there Front cover photograph: Cauldron Snout The Reserve is situated in the heart of © Natural England / Anne Harbron the North Pennines Area of Outstanding Natural Beauty. It is in two parts on either Natural England is here to conserve and side of Cow Green Reservoir. enhance the natural environment, for its intrinsic value, the wellbeing and A limited bus service stops at Bowlees, enjoyment of people and the economic High Force and Cow Green on request. prosperity that it brings. There is no bus service to the Cumbria © Natural England 2009 side of the Reserve. ISBN 978-1-84754-115-1 Catalogue Code: NE146 For information on public transport www.naturalengland.org.uk phone the local Tourist Information Natural England publications are available Centres as accessible pdfs from: www.naturalengland.org.uk/publications Middleton-in-Teesdale: 01833 641001 Should an alternative format of this publication be required, please contact Alston: 01434 382244 our enquiries line for more information: 0845 600 3078 or email Appleby: 017683 51177 [email protected] Alston Road Garage [01833 640213] or Printed on Defra Silk comprising 75% Travel line [0870 6082608] can also help. -

THE SCENIC HIGHLIGHTS of the PENNINE WAY Discover the True Spirit of the North on England’S Original National Trail
ENGLAND’S GREAT WALKING TRAILS | THE PENNINE WAY LIMESTONE AND LEGEND: THE SCENIC HIGHLIGHTS OF THE PENNINE WAY Discover the true spirit of the North on England’s original National Trail Steeped in history and traversing spectacular upland landscapes in some of England’s most popular National ParKs, the Pennine Way is the most iconic of England’s Great WalKing Trails. Stretching for 268 miles (435Km) across England’s wild northern uplands, with a combined ascent that eXceeds the height of Mount Everest, it’s also arguably the toughest. OVERVIEW • Distance: 74 miles/120km • Start/Finish: Skipton/Appleby-in-Westmorland • Number of Days: 8 • Grade: Challenging • Theme: History / Geology • Landscape: Type High Hills & Moorland Opened in 1965, the Pennine Way blazed a trail for public access to some of England’s wildest landscapes – hitherto the sole preserve of a wealthy elite. Conceived by founder member of The Ramblers Tom Stephenson and popularised by the legendary Alfred Wainwright, the full route follows the rocky spine of England – stretching from the hills of the Derbyshire Peak District, through the Yorkshire Dales and onwards through Durham and Northumberland to the Scottish Border. Roughly following the line of the watershed from which great rivers like the Tyne and the Tees, the Lune and the Eden, flow east and west respectively, the bulk of this legendary trail lies at more than 1,000ft/305m above sea level. Our shortened 8-day itinerary explores the striking limestone landscapes of Malham in the Yorkshire Dales before climbing up onto the lonely massif of Cross Fell, where the Lakeland Fells are clearly visible across the Eden Valley. -

THE WHIN SILL Quartz-Dolerite, Dark in Colour
The rock itself is mainly fine to medium-grained THE WHIN SILL quartz-dolerite, dark in colour. It is tough stuff Alan Gill and makes excellent road stone. You may well have driven on it. One of the many interesting geological features in the north of England is the Whin Sill. This is essentially a subterranean layer of igneous rock underlying much of Northumberland, north east Cumbria, along the Pennine escarpment and in Teesdale. Its area is estimated to be at least 5000 sq km. The maximum thickness recorded is 75m but the average is between 25m and 50m. In many places the intrusion separates into two or more layers divided by several hundred metres induced by joint and fault planes. There are dykes associated with the Whin Sill forming together one single petrographic province. It is possible that these were the conduits through which the magma flowed prior to solidification. The proximity of the sill to the Carboniferous Limestone Series led some geologists to conclude that the sill originated as a contemporaneous lava flow. Sedgwick however advocated its Fig 2: High Force on the River Tees showing the intrusive origin as early as 1826, and was proved vertical structure of the Whin Sill resting on the right later in 1870. It was the subject, of pioneer horizontal Tynebottom Limestone. isotopic age determination by Arthur Holmes. The evidence suggests that the Whin Sill was intruded in late Carboniferous Times; an age of 295 +/- 6my has been computed. There are many good outcrops; the sea cliffs of the Farne Islands - the Romans saw a good thing when they built their wall along the north facing escarpments. -

THE TEESDALE WAY About the Author Martin Collins Is a Freelance Author, Photo-Journalist and Cartographer, As Well As a Regular Contributor to the UK Outdoor Media
THE TEESDALE WAY About the author Martin Collins is a freelance author, photo-journalist and cartographer, as well as a regular contributor to the UK outdoor media. First walking the GR5 in 1981 kindled a passion for the French Alps that remains as strong as ever. He has since written over twenty books for walkers embracing the coast, mountains and countryside of the UK and parts of Europe. He has three children, and lives in north Wales on the edge of the Snowdonia National Park. THE TEESDALE WAY by Martin Collins I’ve wandered many a weary mile, And in strange countries been; I’ve dwelt in towns and on wild moors, And curious sights I’ve seen; But still my heart clings to the dale Where Tees rolls to the sea; Compared with what I’ve seen I’ll say The Teesdale hills for me. (Richard Watson, the ‘Teesdale Poet’ – born Middleton-in-Teesdale 1833, died there 1891) JUNIPER HOUSE, MURLEY MOSS, OXENHOLME ROAD, KENDAL, CUMBRIA LA9 7RL www.cicerone.co.uk © Martin Collins 2005 CONTENTS Second edition 2005 ISBN: 978 1 85284 461 5 Overview map ...............................................................................................6–7 Reprinted 2013 (with updates) and 2019 Map key ............................................................................................................7 First edition 1995 Preface to the Second Edition ............................................................................8 Printed by KHL Printing, Singapore. Introduction ......................................................................................................9 -

Cow Green Geotrail
Outer Spread Outer 19/08/2019 16:42 19/08/2019 1 2.indd (NF) Geotrail Green Cow - Pennines North Illustrations: © NPAP/Jed Atkinson | Front cover: Cauldron Snout Cauldron cover: Front | Atkinson NPAP/Jed © Illustrations: expertise and input. and expertise Partnership’s Geopark Advisory Group for their their for Group Advisory Geopark Partnership’s Survey, with thanks given to members of the AONB AONB the of members to given thanks with Survey, Pennines AONB Partnership and British Geological Geological British and Partnership AONB Pennines This publication is based on original work by the North North the by work original on based is publication This contact the reserve base on 01833 622374. 01833 on base reserve the contact grassland and blanket bog. For more information information more For bog. blanket and grassland from hay meadows and juniper woods to limestone limestone to woods juniper and meadows hay from range of upland habitats typical of the North Pennines, Pennines, North the of typical habitats upland of range NNR. This large reserve contains an almost complete complete almost an contains reserve large This NNR. This trail is within the Moor House-Upper Teesdale Teesdale House-Upper Moor the within is trail This northpenninesaonb [email protected] northpennines Moor House-Upper Teesdale National Nature Reserve (NNR) Reserve Nature National Teesdale House-Upper Moor +44 (0)1388 528801 (0)1388 +44 @NorthPennAONB www.northpennines.org.uk heritage, visit www.northpennines.org.uk visit heritage, NorthPenninesAONB North Pennines AONB Partnership AONB Pennines North others and to fi nd out more about our geological geological our about more out nd fi to and others publications for the North Pennines. -

Long-Distance Walks
Long distance walks Page 1 of 47 Long-distance walks Long distance walks Page 2 of 47 Walking has always been a rewarding activity to me. As a boy in Rushden I walked to school, to visit relatives, to Boys Brigade meetings and to the Methodist Church. You walked or cycled. In Cambridge it was often easier to walk than cycle. When I was at home during vacations with Anne living in Irchester, 3 miles away, I often walked back late at night cursing an early last bus. When our boys were young we went to the nearby National Trust properties at Styal or Lyme Park to play and walk. One standout memory is coming down from the Bowstones at Lyme Park with Richard, aged 2, showing signs of hypothermia. And my interest in fungi started during these walks as the boys were ace at spotting them. We still walk most days for 2 or 3 miles although often with a stop for coffee along the way. We know, and can rate, every coffee shop in the area. So it is no surprise that I have some long-distance walks to write about. The first, and longest, was the Pennine Way done as a single walk over 14 days in 1975. It was the first official long distance route created in the UK. Now they criss-cross the country. You are as likely to see a fingerpost saying something like Hatters Way as simply Public Footpath. My next experience – the northern section of the Pembrokeshire Coastal Path – differed in almost every way. -

The Mineralogical Magazine
THE MINERALOGICAL MAGAZINE AND 5OURbT2~L OF THE :~/iI~TERY~LO GIC2~:L SOCIET~ r. No. 122. September, 1928. Vol. XXI. The age and composition of the Whin Sill and the related dikes of the north of .England) (With Plate X.) :By ARTHUR HOL~ES, D.Sc., A.R.C.S., F.G.S. Professor of Geology, The University, Durham, and H. F. HAawooD, M.Sc,, Ph.D., A.I.C. Lecturer in Chemistry, Imperial Co]]ege, London. [Read March 20, 1928.] CONTENTS. 1. Introduction and Literature 494~ 6. The Holy Island Dike- 2. General Survey ...... 496 Echelon ... .... ** 514 3. Petrological Characters of the 7. The High Green Dike~ ~ormal Rocks ...... 500 Echelon ... 516 Felspars ......... 501 8. The St.Oswald'sChapel Dike- Pyroxenes ......... 503 Echelon ......... 520 Note on the Pyroxenes of 9. The Hett Dike-System and Inninmorite .~ ... 507 the Wackerfield Dike ... 523 4. 1KineL:al Composition of the 10. Geological Age of the In- Whin Sill 508 trusions 5~.9 5. Chemical Composition of the 11. The Magma-Type and its Dis- Whin Sill ...... 511 tribution ......... 535 I The chemical analyses and calculations of norms have been made by Dr. Harwood. The field work and petrological investigations have been carried out by Professor Holmes~ who is also responsible for the ~ riting of the paper. L1 494 A, lqOLMES AND lt. F. HAR~V00D ON 1. INTRODUCTIONAND LITERATURE. HE system of intrusive sheets in the Lower Carboniferous fermations T of the north of England, familiar to geologists throughout the world as the Great Whin Sill, has a voluminous early literature which it is unnecessal T to summarize here. -

The Whin Sill You Can See Dramatic Exposures of the Whin Sill at Several Places in Northern England
Where to see the Whin Sill You can see dramatic exposures of the Whin Sill at several places in northern England. Visit Upper Teesdale in the North Pennines to see it at the waterfalls of High Force, Low Force and Cauldron Snout, and at the crags of Holwick Scars, Falcon Clints and Cronkley Scar. The North Pennines is one of England’s most special places – a peaceful, unspoilt landscape with a rich history and vibrant Holwick Scars natural beauty. In recognition of this it is designated as an ▲ Area of Outstanding Natural Beauty (AONB). The area is also a The Whin Sill Global Geopark – an accolade endorsed by UNESCO. ▲ Falcon Clints The Whin Sill is one of the most famous and dramatic natural features of the North Pennines – and its origins are just as spectacular. North Pennines AONB Partnership NorthPenninesAONB www.northpennines.org.uk @NorthPennAONB +44 (0)1388 528801 northpennines [email protected] northpennaonb ▲ On the North Pennine escarpment, near Dufton, the The AONB Partnership has a Green Whin Sill forms a spectacular ring of cliffs towering over Tourism award for its corporate office the deep valley of High Cup Gill. In Weardale you can see the Little Whin Sill in the Rookhope Burn and in the disused quarry at Greenfoot, near Stanhope. In the Northumberland Coast AONB the Whin Sill forms the Farne Islands and some dramatic stretches of coastline. In the Supported by: Northumberland National Park the Whin Sill is a formidable natural rampart for Hadrian’s Wall. Front cover photo: High Force, Upper Teesdale Unless otherwise credited, all photographs © NPAP/Elizabeth Pickett 01/15/8K Printed on 150gsm Revive 100 Silk The Whin Sill is one of the most famous and dramatic natural features of Working the whinstone the North Pennines – and its origins are just as spectacular.