Tracy L. Kivell Pierre Lemelin Brian G. Richmond Daniel Schmitt
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Musculoskeletal Ultrasound Technical Guidelines II. Elbow
European Society of MusculoSkeletal Radiology Musculoskeletal Ultrasound Technical Guidelines II. Elbow Ian Beggs, UK Stefano Bianchi, Switzerland Angel Bueno, Spain Michel Cohen, France Michel Court-Payen, Denmark Andrew Grainger, UK Franz Kainberger, Austria Andrea Klauser, Austria Carlo Martinoli, Italy Eugene McNally, UK Philip J. O’Connor, UK Philippe Peetrons, Belgium Monique Reijnierse, The Netherlands Philipp Remplik, Germany Enzo Silvestri, Italy Elbow Note The systematic scanning technique described below is only theoretical, considering the fact that the examination of the elbow is, for the most, focused to one quadrant only of the joint based on clinical findings. 1 ANTERIOR ELBOW For examination of the anterior elbow, the patient is seated facing the examiner with the elbow in an extension position over the table. The patient is asked to extend the elbow and supinate the fore- arm. A slight bending of the patient’s body toward the examined side makes full supination and as- sessment of the anterior compartment easier. Full elbow extension can be obtained by placing a pillow under the joint. Transverse US images are first obtained by sweeping the probe from approximately 5cm above to 5cm below the trochlea-ulna joint, a Pr perpendicular to the humeral shaft. Cranial US images of the supracondylar region reveal the superficial biceps and the deep brachialis mu- Br scles. Alongside and medial to these muscles, follow the brachial artery and the median nerve: * the nerve lies medially to the artery. * Legend: a, brachial artery; arrow, median nerve; arrowheads, distal biceps tendon; asterisks, articular cartilage of the Humerus humeral trochlea; Br, brachialis muscle; Pr, pronator muscle 2 distal biceps tendon: technique The distal biceps tendon is examined while keeping the patient’s forearm in maximal supination to bring the tendon insertion on the radial tuberosity into view. -

Structure of the Flexor Digitorum Superficialis
Okajimas Folia Anat. Jpn., 56(5) : 277-288, December 1979 Structure of the Flexor Digitorum Superficialis By OSAMU OHTANI Department of Anatomy, Okayama University Medical School, Okayama 700, Japan (Director : Prof. Dr. H. Outi) —Received for Publication, March 9, 1979— Key Words: Flexor digitorum superficialis, Striated muscle, Forearm musculature. Summary. Fifty-two forearms of 26 Japanese adult cadavers were examined. The flexor digitorum superficialis can be separated into two layers: a superficial layer and a deep layer. The superficial layer is composed of the radial head and the superficial part of the humeroulnar head. It forms two muscle bellies which give rise to the tendons for the third and fourth digits, respectively. The deep layer is composed of the deep part of the humeroulnar head. After forming an inter- mediate tendon, the deep layer also divides into two fleshy bellies which give rise to the tendons for the second and fifth digits, respectively. Based on the mode of occurrence of the communicating muscle fasciculi between the superficial layer and the deep layer, the flexor digitorum superficialis can be classified into four types. Type I muscle has no communicating fasciculus (4/52, 7.7%). Type II muscle has a communicating muscle fasciculus between the intermediate tendon and the muscle belly for the fourth digit (muscle fasciculus A) (29/52, 55.8%). Type III muscle has the muscle fasciculus A as well as another communicating muscle fasciculus between the intermediate tendon and the belly for the third digit (muscle fasciculus B) (18/52, 34.6%). Type IV muscle has the muscle fasciculus B only (1/52, 1.9%). -

Disclosures Flexor Pronator Strain
11/19/2018 Flexor Pronator Strain, Epicondylitis, Avulsions Lee Kaplan, MD Director, UHealth Sports Medicine Institute Professor, Department of Orthopaedics, Kinesiology & Sports Science, Biomedical Engineering Medical Director and Head Team Physician, University of Miami Athletics Medical Director and Head Team Physician, Miami Marlins Josue Acevedo, MD Orthopaedics Sports Medicine fellow Disclosures • I have no conflicts of interest in relation to this presentation 2 Flexor Pronator Strain • Anatomy • Function • Forces • Clinical Presentation • Imaging Studies • Treatment 3 1 11/19/2018 Anatomy • Common Flexor Tendon origin - Medial Epicondyle Pronator Teres FCR Palmaris Longus FDS FCU 4 Function • Provides dynamic support to valgus stress during throwing motion • Contraction during early arm acceleration resists valgus and flexes wrist during ball release 5 Elbow Forces • Elbow valgus torque peaks at late cocking phase - during maximal shoulder external rotation (MER) • Can approach torque up to 120 N.m • Significantly higher torque in pitchers who enduring an injury Sawbick et al. JSES, 2004 Adam WA. AJSM, 2010. 6 2 11/19/2018 • Fastballs caused the greatest torque • Curveballs produced the greatest arm speed . • Predictors of increased elbow torque • Increased ball velocity • Higher BMI • Decreased arm slot • Protectors against elbow torque. • Increasing age • longer arm length • greater elbow circumference were independent 7 • VAS fatigue scores increased 0.72 points per inning • Medial elbow torque increased beyond inning 3, increase of 0.84 N·m each inning. • Pitch velocity decreased (0.28 mph per inning) • There were no differences in arm rotation or arm speed as the game progressed. • Arm slot decreased with each successive inning (0.73° on average per inning) ***These findings signify a possible relationship between fatigue and injury risk. -

Old World Monkeys
OLD WORLD MONKEYS Edited by Paul F. Whitehead and Clifford J. Jolly The Pitt Building, Trumpington Street, Cambridge CB2 1RP, United Kingdom The Edinburgh Building, Cambridge CB2 2RU, UK http://www.cup.cam.ac.uk 40 West 20th Street, New York, NY 10011-4211, USA http://www.cup.org 10 Stamford Road, Oakleigh, Melbourne 3166, Australia Ruiz de Alarco´n 13, 28014 Madrid, Spain © Cambridge University Press 2000 This book is in copyright. Subject to statutory exception and to the provisions of relevant collective licensing agreements, no reproduction of any part may take place without the written permission of Cambridge University Press. First published 2000 Printed in the United Kingdom at the University Press, Cambridge Typeface Times NR 10/13pt. System QuarkXPress® [] A catalogue record for this book is available from the British Library Library of Congress Cataloguing in Publication data Old world monkeys / edited by Paul F. Whitehead & Clifford J. Jolly. p. cm. ISBN 0 521 57124 3 (hardcover) 1. Cercopithecidae. I. Whitehead, Paul F. (Paul Frederick), 1954– . II. Jolly, Clifford J., 1939– . QL737.P930545 2000 599.8Ј6–dc21 99-20192 CIP ISBN 0 521 57124 3 hardback Contents List of contributors page vii Preface x 1 Old World monkeys: three decades of development and change in the study of the Cercopithecoidea Clifford J. Jolly and Paul F. Whitehead 1 2 The molecular systematics of the Cercopithecidae Todd R. Disotell 29 3 Molecular genetic variation and population structure in Papio baboons Jeffrey Rogers 57 4 The phylogeny of the Cercopithecoidea Colin P. Groves 77 5 Ontogeny of the nasal capsule in cercopithecoids: a contribution to the comparative and evolutionary morphology of catarrhines Wolfgang Maier 99 6 Old World monkey origins and diversification: an evolutionary study of diet and dentition Brenda R. -

Anatomical Considerations of Fascial Release in Ulnar Nerve Transposition: a Concept Revisited
LABORATORY INVESTIGATION J Neurosurg 123:1216–1222, 2015 Anatomical considerations of fascial release in ulnar nerve transposition: a concept revisited Mark A. Mahan, MD,1 Jaime Gasco, MD,2 David B. Mokhtee, MD,3 and Justin M. Brown, MD4 1Division of Neurological Surgery, Barrow Neurological Institute, St. Joseph’s Hospital and Medical Center, Phoenix, Arizona; 2Division of Neurological Surgery, University of Texas Medical Branch, Galveston, Texas; 3Tulsa Bone and Joint Associates, Tulsa, Oklahoma; and 4Division of Neurosurgery, University of California, San Diego, La Jolla, California OBJECT Surgical transposition of the ulnar nerve to alleviate entrapment may cause otherwise normal structures to become new sources of nerve compression. Recurrent or persistent neuropathy after anterior transposition is commonly attributable to a new distal compression. The authors sought to clarify the anatomical relationship of the ulnar nerve to the common aponeurosis of the humeral head of the flexor carpi ulnaris (FCU) and flexor digitorum superficialis (FDS) muscles following anterior transposition of the nerve. METHODS The intermuscular septa of the proximal forearm were explored in 26 fresh cadaveric specimens. The fibrous septa and common aponeurotic insertions of the flexor-pronator muscle mass were evaluated in relation to the ulnar nerve, with particular attention to the effect of transposition upon the nerve in this region. RESULTS An intermuscular aponeurosis associated with the FCU and FDS muscles was present in all specimens. Transposition consistently resulted in angulation of the nerve during elbow flexion when this fascial septum was not released. The proximal site at which the nerve began to traverse this fascial structure was found to be an average of 3.9 cm (SD 0.7 cm) from the medial epicondyle. -

Areas 1- Ern Africa
Kroeber Anthropological Society Papers, Nos. 71-72, 1990 Diet, Species Diversity and Distribution of African Fossil Baboons Brenda R. Benefit and Monte L. McCrossin Based on measurements ofmolarfeatures shown to befunctionally correlated with the proportions of fruits and leaves in the diets ofextant monkeys, Plio-Pleistocenepapionin baboonsfrom southern Africa are shown to have included more herbaceous resources in their diets and to have exploited more open country habitats than did the highlyfrugivorousforest dwelling eastern African species. The diets ofall species offossil Theropithecus are reconstructed to have included morefruits than the diets ofextant Theropithecus gelada. Theropithecus brumpti, T. quadratirostris and T. darti have greater capacitiesfor shearing, thinner enamel and less emphases on the transverse component ofmastication than T. oswaldi, and are therefore interpreted to have consumed leaves rather than grass. Since these species are more ancient than the grass-eating, more open country dwelling T. oswaldi, the origin ofthe genus Thero- pithecus is attributed tofolivorous adaptations by largepapionins inforest environments rather than to savannah adapted grass-eaters. Reconstructions ofdiet and habitat are used to explain differences in the relative abundance and diversity offossil baboons in eastern andsouthern Africa. INTRODUCTION abundance between eastern and southern Africa is observed for members of the Papionina (Papio, Interpretations of the dietary habits of fossil Cercocebus, Parapapio, Gorgopithecus, and Old World monkeys have been based largely on Dinopithecus). [We follow Szalay and Delson analogies to extant mammals with lophodont teeth (1979) in recognizing two tribes of cercopithe- (Jolly 1970; Napier 1970; Delson 1975; Andrews cines, Cercopithecini and Papionini, and three 1981; Andrews and Aiello 1984; Temerin and subtribes of the Papionini: Theropithecina (gela- Cant 1983). -

Les 1000 Premiers Jours De Vie Dans Les Populations Du Présent Et Du Passé the First 1,000 Days of Life in Past and Present Populations
Bulletins et mémoires de la Société d’Anthropologie de Paris 33 (1) | 2021 Les 1000 premiers jours de vie dans les populations du présent et du passé The first 1,000 days of life in past and present populations Fernando Ramirez Rozzi, Gwenaëlle Goude, Estelle Herrscher, François Marchal et Aline Thomas (dir.) Édition électronique URL : https://journals.openedition.org/bmsap/6913 DOI : 10.4000/bmsap.6913 ISSN : 1777-5469 Éditeur Société d'Anthropologie de Paris Référence électronique Fernando Ramirez Rozzi, Gwenaëlle Goude, Estelle Herrscher, François Marchal et Aline Thomas (dir.), Bulletins et mémoires de la Société d’Anthropologie de Paris, 33 (1) | 2021, « Les 1000 premiers jours de vie dans les populations du présent et du passé » [En ligne], mis en ligne le 13 mars 2021, consulté le 03 juin 2021. URL : https://journals.openedition.org/bmsap/6913 ; DOI : https://doi.org/10.4000/ bmsap.6913 Description de couverture La forme du bassin humain sous influence de forces sélectives locales Mitteroecker et al., 2021 (cliché : Magdalena FISCHER) Les contenus des Bulletins et mémoires de la Société d’Anthropologie de Paris sont mis à disposition selon les termes de la licence Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License. BMSAP Bulletins et Mémoires de la Société d’Anthropologie de Paris Éditions Les Bulletins et Mémoires de la Société d’Anthropologie de Paris temps que la Société d’Anthropologie de Paris (SAP), en 1859, et sont ainsi la plus ancienne Société d’anthropologie de Paris publication au monde dans le domaine de l’anthropologie biologique. (BMSAP) ont été créés en même BMSAP 17 Place du Trocadéro et du 11 Novembre 75016Musée Paris,de l’Homme France Les BMSAP couvrent, de manière pluridisciplinaire, les divers champs de l’anthropologie, aux Directeur de publication frontières du biologique et du culturel. -

Phylogenetic Relationships of Living and Fossil African Papionins: Combined Evidence from Morphology and Molecules
City University of New York (CUNY) CUNY Academic Works Publications and Research Hunter College 2018 Phylogenetic relationships of living and fossil African papionins: Combined evidence from morphology and molecules Kelsey D. Pugh The Graduate Center, City University of New York Christopher C. Gilbert CUNY Hunter College How does access to this work benefit ou?y Let us know! More information about this work at: https://academicworks.cuny.edu/hc_pubs/647 Discover additional works at: https://academicworks.cuny.edu This work is made publicly available by the City University of New York (CUNY). Contact: [email protected] Journal of Human Evolution 123 (2018) 35e51 Contents lists available at ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol Phylogenetic relationships of living and fossil African papionins: Combined evidence from morphology and molecules * Kelsey D. Pugh a, b, , Christopher C. Gilbert a, b, c a PhD Program in Anthropology, Graduate Center of the City University of New York, 365 Fifth Avenue, New York, NY 10016, USA b New York Consortium in Evolutionary Primatology (NYCEP), USA c Department of Anthropology, Hunter College of the City University of New York, 695 Park Avenue, New York, NY 10065, USA article info abstract Article history: African papionins are a highly successful subtribe of Old World monkeys with an extensive fossil record. Received 7 November 2017 On the basis of both molecular and morphological data, crown African papionins are divided into two Accepted 1 June 2018 clades: Cercocebus/Mandrillus and Papio/Lophocebus/Rungwecebus/Theropithecus (P/L/R/T), though Available online 26 July 2018 phylogenetic relationships in the latter clade, among both fossil and extant taxa, remain difficult to resolve. -

Partial Skeleton of Theropithecus Brumpti (Primates, Cercopithecidae) from the Chemeron Formation of the Tugen Hills, Kenya
Journal of Human Evolution 61 (2011) 347e362 Contents lists available at ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol Partial skeleton of Theropithecus brumpti (Primates, Cercopithecidae) from the Chemeron Formation of the Tugen Hills, Kenya Christopher C. Gilbert a,b,c,*, Emily D. Goble d, John D. Kingston e, Andrew Hill d a Department of Anthropology, Hunter College of the City University of New York, 695 Park Avenue, NY 10021, USA b Department of Anthropology, Graduate Center of the City University of New York, 365 Fifth Avenue, NY 10016, USA c New York Consortium in Evolutionary Primatology, New York, NY, USA d Department of Anthropology, Yale University, PO Box 208277, New Haven, CT 06520-8277, USA e Department of Anthropology, Emory University, 1557 Dickey Drive, Atlanta, GA 30322, USA article info abstract Article history: Here we describe a complete skull and partial skeleton of a large cercopithecoid monkey (KNM-TH Received 27 October 2010 46700) discovered in the Chemeron Formation of the Tugen Hills at BPRP Site #152 (2.63 Ma). Associated Accepted 21 April 2011 with the skeleton was a mandible of an infant cercopithecoid (KNM-TH 48364), also described here. KNM-TH 46700 represents an aged adult female of Theropithecus brumpti, a successful Pliocene papionin Keywords: taxon better known from the Omo Shungura Formation in Ethiopia and sites east and west of Lake Cercopithecoid Turkana, Kenya. While the morphology of male T. brumpti is well-documented, including a partial Papionin skeleton with both cranial and postcranial material, the female T. brumpti morphotype is not well- Crania fi Postcrania known. -
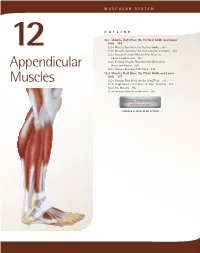
Appendicular Muscles 355
MUSCULAR SYSTEM OUTLINE 12.1 Muscles That Move the Pectoral Girdle and Upper Limb 355 12.1a Muscles That Move the Pectoral Girdle 355 12 12.1b Muscles That Move the Glenohumeral Joint/Arm 360 12.1c Arm and Forearm Muscles That Move the Elbow Joint/Forearm 363 12.1d Forearm Muscles That Move the Wrist Joint, Appendicular Hand, and Fingers 366 12.1e Intrinsic Muscles of the Hand 374 12.2 Muscles That Move the Pelvic Girdle and Lower Limb 377 Muscles 12.2a Muscles That Move the Hip Joint/Thigh 377 12.2b Thigh Muscles That Move the Knee Joint/Leg 381 12.2c Leg Muscles 385 12.2d Intrinsic Muscles of the Foot 391 MODULE 6: MUSCULAR SYSTEM mck78097_ch12_354-396.indd 354 2/14/11 3:25 PM Chapter Twelve Appendicular Muscles 355 he appendicular muscles control the movements of the upper 2. Identify the muscles that move the scapula and their actions. T and lower limbs, and stabilize and control the movements 3. Name the muscles of the glenohumeral joint, and explain of the pectoral and pelvic girdles. These muscles are organized how each moves the humerus. into groups based on their location in the body or the part of 4. Locate and name the muscles that move the elbow joint. the skeleton they move. Beyond their individual activities, these 5. Identify the muscles of the forearm, wrist joint, fingers, muscles also work in groups that are either synergistic or antago- and thumb. nistic. Refer to figure 10.14 to review how muscles are named, and Muscles that move the pectoral girdle and upper limbs are recall the first Study Tip! from chapter 11 that gives suggestions organized into specific groups: (1) muscles that move the pectoral for learning the muscles. -

Diet of Theropithecus from 4 to 1 Ma in Kenya
Diet of Theropithecus from 4 to 1 Ma in Kenya Thure E. Cerlinga,b,1, Kendra L. Chritzb, Nina G. Jablonskic, Meave G. Leakeyd,e,f, and Fredrick Kyalo Manthif Departments of aGeology and Geophysics and bBiology, University of Utah, Salt Lake City, UT 84013; cDepartment of Anthropology, Penn State, University Park, PA 16802; dTurkana Basin Institute, 00502 Nairobi, Kenya; eDepartment of Anthropology, Stony Brook University, Stony Brook, NY 11794; and fNational Museums of Kenya, 00100 Nairobi, Kenya Edited by James O’Connell, University of Utah, Salt Lake City, UT, and approved April 15, 2013 (received for review December 23, 2012) Theropithecus was a common large-bodied primate that co- grass as effectively as an equid, and they can also ferment cellular occurred with hominins in many Plio-Pleistocene deposits in East material from grass in their hindguts but less effectively than and South Africa. Stable isotope analyses of tooth enamel from T. a zebra, which may have aided in this adaptation (6, 12, 13). Even brumpti (4.0–2.5 Ma) and T. oswaldi (2.0–1.0 Ma) in Kenya show with the richness of the genus’s fossil record and the many pa- that the earliest Theropithecus at 4 Ma had a diet dominated by C4 leoecological and functional anatomical studies that have spec- resources. Progressively, this genus increased the proportion of ulated on the respective habitat and dietary preferences of the C4-derived resources in its diet and by 1.0 Ma, had a diet that was T. brumpti and T. oswaldi lineages, many questions remain about nearly 100% C4-derived. -
Development, Function and Evolution of Teeth
Development, Function and Evolution of Teeth Over the past 20 years there has been an explosion of information generated by scienti®c research. One of the bene®ciaries of this has been the study of morphology, where new techniques and analyses have led to insights into a wide range of topics. Advances in genetics, histology, microstructure, biomechanics and morphometrics have allowed researchers to view teeth from new perspectives. However, up to now there has been little communication between researchers in the different ®elds of dental research. This book brings together for the ®rst time overviews of a wide range of dental topics, linking genes, molecules and developmental mechanisms within an evolutionary framework. Written by leading experts in the ®eld, this book will stimulate co-operative research in ®elds as diverse as palaeontology, molecular biology, developmental biology and functional morphology. MARK F.TEAFORD is Professor of Anatomy at the Johns Hopkins University School of Medicine. MOYA MEREDITH SMITH is Professor of Evolutionary Dentoskeletal Biology at the University of London, and is based at the Dental Institute of King's College London, Guy's Campus. MARK W. J. FERGUSON is Professor of Basic Dental Sciences in the School of Biological Sciences at the University of Manchester. Development, Function and Evolution of Teeth Edited by Mark F. Teaford, Moya Meredith Smith and Mark W. J. Ferguson Cambridge, New York, Melbourne, Madrid, Cape Town, Singapore, São Paulo Cambridge University Press The Edinburgh Building, Cambridge , United Kingdom Published in the United States by Cambridge University Press, New York www.cambridge.org Information on this title: www.cambridge.org/9780521570114 © Cambridge University Press 2000 This book is in copyright.