A Brief History of Whale Evolution: As Supported by The
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
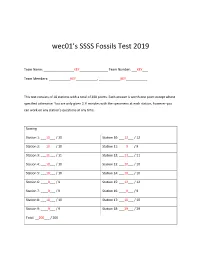
Wec01's SSSS Fossils Test 2019
wec01’s SSSS Fossils Test 2019 Team Name: _________________KEY________________ Team Number: ___KEY___ Team Members: ____________KEY____________, ____________KEY____________ This test consists of 18 stations with a total of 200 points. Each answer is worth one point except where specified otherwise. You are only given 2 ½ minutes with the specimens at each station, however you can work on any station’s questions at any time. Scoring Station 1: ___10___ / 10 Station 10: ___12___ / 12 Station 2: ___10___ / 10 Station 11: ____9___ / 9 Station 3: ___11___ / 11 Station 12: ___11___ / 11 Station 4: ___10___ / 10 Station 13: ___10___ / 10 Station 5: ___10___ / 10 Station 14: ___10___ / 10 Station 6: ____9___ / 9 Station 15: ___12___ / 12 Station 7: ____9___ / 9 Station 16: ____9___ / 9 Station 8: ___10___ / 10 Station 17: ___10___ / 10 Station 9: ____9___ / 9 Station 18: ___29___ / 29 Total: __200___ / 200 Team Number: _KEY_ Station 1: Dinosaurs (10 pt) 1. Identify the genus of specimen A Tyrannosaurus (1 pt) 2. Identify the genus of specimen B Stegosaurus (1 pt) 3. Identify the genus of specimen C Allosaurus (1 pt) 4. Which specimen(s) (A, B, or C) are A, C (1 pt) Saurischians? 5. Which two specimens (A, B, or C) lived at B, C (1 pt) the same time? 6. Identify the genus of specimen D Velociraptor (1 pt) 7. Identify the genus of specimen E Coelophysis (1 pt) 8. Which specimen (D or E) is commonly E (1 pt) found in Ghost Ranch, New Mexico? 9. Which specimen (A, B, C, D, or E) would D (1 pt) specimen F have been found on? 10. -

PDF of Manuscript and Figures
The triple origin of whales DAVID PETERS Independent Researcher 311 Collinsville Avenue, Collinsville, IL 62234, USA [email protected] 314-323-7776 July 13, 2018 RH: PETERS—TRIPLE ORIGIN OF WHALES Keywords: Cetacea, Mysticeti, Odontoceti, Phylogenetic analyis ABSTRACT—Workers presume the traditional whale clade, Cetacea, is monophyletic when they support a hypothesis of relationships for baleen whales (Mysticeti) rooted on stem members of the toothed whale clade (Odontoceti). Here a wider gamut phylogenetic analysis recovers Archaeoceti + Odontoceti far apart from Mysticeti and right whales apart from other mysticetes. The three whale clades had semi-aquatic ancestors with four limbs. The clade Odontoceti arises from a lineage that includes archaeocetids, pakicetids, tenrecs, elephant shrews and anagalids: all predators. The clade Mysticeti arises from a lineage that includes desmostylians, anthracobunids, cambaytheres, hippos and mesonychids: none predators. Right whales are derived from a sister to Desmostylus. Other mysticetes arise from a sister to the RBCM specimen attributed to Behemotops. Basal mysticetes include Caperea (for right whales) and Miocaperea (for all other mysticetes). Cetotheres are not related to aetiocetids. Whales and hippos are not related to artiodactyls. Rather the artiodactyl-type ankle found in basal archaeocetes is also found in the tenrec/odontocete clade. Former mesonychids, Sinonyx and Andrewsarchus, nest close to tenrecs. These are novel observations and hypotheses of mammal interrelationships based on morphology and a wide gamut taxon list that includes relevant taxa that prior studies ignored. Here some taxa are tested together for the first time, so they nest together for the first time. INTRODUCTION Marx and Fordyce (2015) reported the genesis of the baleen whale clade (Mysticeti) extended back to Zygorhiza, Physeter and other toothed whales (Archaeoceti + Odontoceti). -

High School Living Earth Evidence for Evolution Lessons Name: School: Teacher
DO NOT EDIT--Changes must be made through “File info” CorrectionKey=CA-B High School Living Earth Evidence for Evolution Lessons Name: School: Teacher: The Unit should take approximately 4 days complete. Read each section and complete the tasks. DO NOT EDIT--Changes must be made through “File info” CorrectionKey=CA-B FIGURE 1: This creosote ring in the Mojave Desert is estimated to be 11 700 years old . This makes it one of the oldest living organisms on Earth . The creosote bush is thought to be the most drought-tolerant plant in North America. It has a variety of adaptations to its desert environment, including its reproductive tendency to clone outward in rings rather than rely solely on seed production. The plant’s leaves are coated in a foul-tasting resin that protects it from water loss through evaporation and from grazing. It only opens its stomata in the morning to pull in carbon dioxide for photosynthesis from the more humid air and closes them as the day’s temperature increases. It also has a root system that consists of both an exceptionally long tap root and a vast network of shallow feeder roots. Creosote bushes exhibit two different shapes to fit different microclimates. In drier areas, the plant has a cone shape in which stems funnel rainwater into the taproot. In wetter areas, the bush has a more rounded shape that provides shade to its shallow feeder roots. PREDICT How do species change over time to adjust to varying conditions? DRIVING QUESTIONS As you move through the unit, gather evidence to help you answer the following questions. -

Whale Evolution Lab – Page 1
Whale Evolution Lab – Page 1 Whale Evolution Lab Adapted from http://www.indiana.edu/~ensiweb PRELAB (recommended)… FROM LAND TO WATER: A Whale Evolution Internet Activity http://www.indiana.edu/~ensiweb/lessons/whalekiosk.html Click on “SCREEN” to “Find Out More” A. Anatomy B. Fossil record C. Molecular evidence A. WHALE ANATOMY 1. What does the Latin word “cetus” mean? 2. What three groups of organisms are considered cetaceans? 3. What are the two subgroups of cetaceans? 4. What characterizes the subgroup Odontoceti? 5. What characterizes the subgroup Mysticeti? 6. Follow the instructions given to compare anatomical parts. Click on the labels to compare the whale’s anatomy with that of a fish and a cat. Fill out the chart below with your answers to each structure, by placing an “X” under the organism whose structure is more similar to the whale’s. Structure FISH CAT Structure FISH CAT Ears Forelimb Eyes Jaw Lungs Mammary gland 7. According to the anatomical evidence, which organism is more closely related to a whale…fish or cat? 8. Draw and label the cladogram that you created for the whale, fish, and cat below. 9. What is the relationship between whales and cats? B. FOSSIL RECORD 1. What is a fossil? 2. What are the most likely parts to become fossilized? 3. What are trace fossils? List some examples. 4. What is a coprolite? 5. What fossilized anatomical structure can be useful to anatomists? 6. Compare the fossil teeth of whales to the other organisms on the website. What sort of organism has fossil teeth most similar to whale teeth? 7. -

Thomas Jefferson Meg Tooth
The ECPHORA The Newsletter of the Calvert Marine Museum Fossil Club Volume 30 Number 3 September 2015 Thomas Jefferson Meg Tooth Features Thomas Jefferson Meg The catalogue number Review; Walking is: ANSP 959 Whales Inside The tooth came from Ricehope Estate, Snaggletooth Shark Cooper River, Exhibit South Carolina. Tiktaalik Clavatulidae In 1806, it was Juvenile Bald Eagle originally collected or Sculpting Whale Shark owned by Dr. William Moroccan Fossils Reid. Prints in the Sahara Volunteer Outing to Miocene-Pliocene National Geographic coastal plain sediments. Dolphins in the Chesapeake Sloth Tooth Found SharkFest Shark Iconography in Pre-Columbian Panama Hippo Skulls CT- Scanned Squalus sp. Teeth Sperm Whale Teeth On a recent trip to the Academy of Natural Sciences of Drexel University (Philadelphia), Collections Manager Ned Gilmore gave John Nance and me a behind -the-scenes highlights tour. Among the fossils that belonged to Thomas☼ Jefferson (left; American Founding Father, principal author of the Declaration of Independence, and third President of the United States) was this Carcharocles megalodon tooth. Jefferson’s interests and knowledge were encyclopedic; a delight to know that they included paleontology. Hand by J. Nance. Photo by S. Godfrey. Jefferson portrait from: http://www.biography.com/people/thomas-jefferson-9353715 ☼ CALVERT MARINE MUSEUM www.calvertmarinemuseum.com 2 The Ecphora September 2015 Book Review: The Walking 41 million years ago and has worldwide distribution. It was fully aquatic, although it did have residual Whales hind limbs. In later chapters, Professor Thewissen George F. Klein discusses limb development and various genetic factors that make whales, whales. This is a The full title of this book is The Walking complicated topic, but I found these chapters very Whales — From Land to Water in Eight Million clear and readable. -

Intervertebral and Epiphyseal Fusion in the Postnatal Ontogeny of Cetaceans and Terrestrial Mammals
J Mammal Evol DOI 10.1007/s10914-014-9256-7 ORIGINAL PAPER Intervertebral and Epiphyseal Fusion in the Postnatal Ontogeny of Cetaceans and Terrestrial Mammals Meghan M. Moran & Sunil Bajpai & J. Craig George & Robert Suydam & Sharon Usip & J. G. M. Thewissen # Springer Science+Business Media New York 2014 Abstract In this paper we studied three related aspects of the Introduction ontogeny of the vertebral centrum of cetaceans and terrestrial mammals in an evolutionary context. We determined patterns The vertebral column provides support for the body and of ontogenetic fusion of the vertebral epiphyses in bowhead allows for flexibility and mobility (Gegenbaur and Bell whale (Balaena mysticetus) and beluga whale 1878;Hristovaetal.2011; Bruggeman et al. 2012). To (Delphinapterus leucas), comparing those to terrestrial mam- achieve this mobility, individual vertebrae articulate with each mals and Eocene cetaceans. We found that epiphyseal fusion other through cartilaginous intervertebral joints between the is initiated in the neck and the sacral region of terrestrial centra and synovial joints between the pre- and post- mammals, while in recent aquatic mammals epiphyseal fusion zygapophyses. The mobility of each vertebral joint varies is initiated in the neck and caudal regions, suggesting loco- greatly between species as well as along the vertebral column motor pattern and environment affect fusion pattern. We also within a single species. Vertebral column mobility greatly studied bony fusion of the sacrum and evaluated criteria used impacts locomotor style, whether the animal is terrestrial or to homologize cetacean vertebrae with the fused sacrum of aquatic. In aquatic Cetacea, buoyancy counteracts gravity, and terrestrial mammals. We found that the initial ossification of the tail is the main propulsive organ (Fish 1996;Fishetal. -

Constraints on the Timescale of Animal Evolutionary History
Palaeontologia Electronica palaeo-electronica.org Constraints on the timescale of animal evolutionary history Michael J. Benton, Philip C.J. Donoghue, Robert J. Asher, Matt Friedman, Thomas J. Near, and Jakob Vinther ABSTRACT Dating the tree of life is a core endeavor in evolutionary biology. Rates of evolution are fundamental to nearly every evolutionary model and process. Rates need dates. There is much debate on the most appropriate and reasonable ways in which to date the tree of life, and recent work has highlighted some confusions and complexities that can be avoided. Whether phylogenetic trees are dated after they have been estab- lished, or as part of the process of tree finding, practitioners need to know which cali- brations to use. We emphasize the importance of identifying crown (not stem) fossils, levels of confidence in their attribution to the crown, current chronostratigraphic preci- sion, the primacy of the host geological formation and asymmetric confidence intervals. Here we present calibrations for 88 key nodes across the phylogeny of animals, rang- ing from the root of Metazoa to the last common ancestor of Homo sapiens. Close attention to detail is constantly required: for example, the classic bird-mammal date (base of crown Amniota) has often been given as 310-315 Ma; the 2014 international time scale indicates a minimum age of 318 Ma. Michael J. Benton. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Philip C.J. Donoghue. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Robert J. -

Functional Morphology of the Vertebral Column in Remingtonocetus (Mammalia, Cetacea) and the Evolution of Aquatic Locomotion in Early Archaeocetes
Functional Morphology of the Vertebral Column in Remingtonocetus (Mammalia, Cetacea) and the Evolution of Aquatic Locomotion in Early Archaeocetes by Ryan Matthew Bebej A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy (Ecology and Evolutionary Biology) in The University of Michigan 2011 Doctoral Committee: Professor Philip D. Gingerich, Co-Chair Professor Philip Myers, Co-Chair Professor Daniel C. Fisher Professor Paul W. Webb © Ryan Matthew Bebej 2011 To my wonderful wife Melissa, for her infinite love and support ii Acknowledgments First, I would like to thank each of my committee members. I will be forever grateful to my primary mentor, Philip D. Gingerich, for providing me the opportunity of a lifetime, studying the very organisms that sparked my interest in evolution and paleontology in the first place. His encouragement, patience, instruction, and advice have been instrumental in my development as a scholar, and his dedication to his craft has instilled in me the importance of doing careful and solid research. I am extremely grateful to Philip Myers, who graciously consented to be my co-advisor and co-chair early in my career and guided me through some of the most stressful aspects of life as a Ph.D. student (e.g., preliminary examinations). I also thank Paul W. Webb, for his novel thoughts about living in and moving through water, and Daniel C. Fisher, for his insights into functional morphology, 3D modeling, and mammalian paleobiology. My research was almost entirely predicated on cetacean fossils collected through a collaboration of the University of Michigan and the Geological Survey of Pakistan before my arrival in Ann Arbor. -

Thewissen Et Al. Reply Replying To: J
NATURE | Vol 458 | 19 March 2009 BRIEF COMMUNICATIONS ARISING Hippopotamus and whale phylogeny Arising from: J. G. M. Thewissen, L. N. Cooper, M. T. Clementz, S. Bajpai & B. N. Tiwari Nature 450, 1190–1194 (2007) Thewissen etal.1 describe new fossils from India that apparentlysupport fossils, Raoellidae or the raoellid Indohyus is more closely related to a phylogeny that places Cetacea (that is, whales, dolphins, porpoises) as Cetacea than is Hippopotamidae (Fig. 1). Hippopotamidae is the the sister group to the extinct family Raoellidae, and Hippopotamidae exclusive sister group to Cetacea plus Raoellidae in the analysis that as more closely related to pigs and peccaries (that is, Suina) than to down-weights homoplastic characters, althoughin the equallyweighted cetaceans. However, our reanalysis of a modified version of the data set analysis, another topology was equally parsimonious. In that topology, they used2 differs in retaining molecular characters and demonstrates Hippopotamidae moved one node out, being the sister group to an that Hippopotamidae is the closest extant family to Cetacea and that Andrewsarchus, Raoellidae and Cetacea clade. In neither analysis is raoellids are the closest extinct group, consistent with previous phylo- Hippopotamidae closer to the pigs and peccaries than to Cetacea, the genetic studies2,3. This topology supports the view that the aquatic result obtained by Thewissen et al.1. In all our analyses, pachyostosis adaptations in hippopotamids and cetaceans are inherited from their (thickening) of limb bones and bottom walking, which occur in hippo- common ancestor4. potamids9,10, are interpreted to have evolved before the pachyostosis of To conduct our analyses, we started with the same published matrix the auditory bulla, as seen in raoellids and cetaceans1. -

Contributions from the Museum of Paleontology
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN Vol. 25, No. 6, p. 117-124 (2 text-figs.; 1 plate) January 26,1979 CHORLAKKZA HASSANZ, A NEW MIDDLE EOCENE DICHOBUNID (MAMMALIA, ARTIODACTYLA) FROM THE KULDANA FORMATION OF KOHAT (PAKISTAN) PHILIP D. GINGERICH, DONALD E. RUSSELL, DENISE SIGOGNEAU-RUSSELL, AND J.-L. HARTENBERGER MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN ANN ARBOR CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY Gerald R. Smith, Director Robert V. Kesling, Editor Diane Wurzinger, Editor for this number The series of contributions from the Museum of Paleontology is a medium for the publication of papers based chiefly upon the collection in the Museum. When the num- ber of pages issued is sufficient to make a volume, a title page and a table of contents will be sent to libraries on the mailing list, and to individuals upon request. A list of the separate papers may also be obtained. Correspondence should be directed to the Museum of Paleontology, The University of Michigan, Ann Arbor, Michigan, 48109. VOLS. 11-XXV. Parts of volumes may be obtained if available. Price lists available upon inquiry. CHORLAKKLl HASSANZ, A NEW MIDDLE EOCENE DICHOBUNID (MAMMALIA, ARTIODACTYLA) FROM THE KULDANA FORMATION OF KOHAT (PAKISTAN) Philip D. Gingerich' ,Donald E. c us sell^, Denise Sigogneau-Russell2,and J.-L. Hartenberger3 Abstract.- A new genus and species of artiodactyl, Chorlakkia hassani, is de- scribed from the middle Eocene Kuldana Formation in the Kohat District of Pakistan. This is the smallest artiodactyl described from the Paleogene of Asia, and it is one of the smallest artiodactyls yet known. -

Transitional Fossils by Greg
Transitional Fossils By Greg Neyman © Answers In Creation Answers In Creation Website www.answersincreation.org/transitional_fossils.htm Transitional fossils, or the supposed lack thereof, has been used for many years by anti-evolutionists to argue against evolution. Here, I will explain what a transitional fossil is, and why it is not valid as an argument against evolution. A transitional fossil shows the evolutionary development from one species to another. For example, if organism 1 existed 70 million years ago, and organism 2 shows up in the fossil record 5 million years later, then theoretically there should be intermediate species in this 5 million year gap, which shows gradual progression from one species to another. The lack of these "transitional" fossils is proof to young earth creationists that evolution is false. Evolutionists have shown that indeed there are transitional fossils, and there are plenty of examples of them. For instance, see http://www.talkorigins.org/faqs/faq- transitional.html . Here is the key point...even if young earth creationists accept these examples of transitional fossils, they will still claim that there are no transitional fossils! These fossils will be called either unique species, or they will come up with some reason (disease, birth defect, etc) that accounts for the apparent transition feature. Naturally, they will say, "Where are the transitional fossils between these transitional fossils?" If we had a clear fossil record, showing progression every 10,000 years for millions of years, they will not believe it, and will want the "transitional" fossils for the missing 10,000 year period. No amount of evidence will convict them that their belief is wrong. -

Transition of Eocene Whales from Land to Sea: Evidence from Bone Microstructure
RESEARCH ARTICLE Transition of Eocene Whales from Land to Sea: Evidence from Bone Microstructure Alexandra Houssaye1,2*, Paul Tafforeau3, Christian de Muizon4, Philip D. Gingerich5 1 UMR 7179 CNRS/Muséum National d’Histoire Naturelle, Département Ecologie et Gestion de la Biodiversité, Paris, France, 2 Steinmann Institut für Geologie, Paläontologie und Mineralogie, Universität Bonn, Bonn, Germany, 3 European Synchrotron Radiation Facility, Grenoble, France, 4 Sorbonne Universités, CR2P—CNRS, MNHN, UPMC-Paris 6, Département Histoire de la Terre, Muséum National d’Histoire Naturelle, Paris, France, 5 Department of Earth and Environmental Sciences and Museum of Paleontology, University of Michigan, Ann Arbor, Michigan, United States of America a11111 * [email protected] Abstract Cetacea are secondarily aquatic amniotes that underwent their land-to-sea transition during OPEN ACCESS the Eocene. Primitive forms, called archaeocetes, include five families with distinct degrees Citation: Houssaye A, Tafforeau P, de Muizon C, of adaptation to an aquatic life, swimming mode and abilities that remain difficult to estimate. Gingerich PD (2015) Transition of Eocene Whales The lifestyle of early cetaceans is investigated by analysis of microanatomical features in from Land to Sea: Evidence from Bone postcranial elements of archaeocetes. We document the internal structure of long bones, Microstructure. PLoS ONE 10(2): e0118409. ribs and vertebrae in fifteen specimens belonging to the three more derived archaeocete doi:10.1371/journal.pone.0118409 families — Remingtonocetidae, Protocetidae, and Basilosauridae — using microtomogra- Academic Editor: Brian Lee Beatty, New York phy and virtual thin-sectioning. This enables us to discuss the osseous specializations ob- Institute of Technology College of Osteopathic Medicine, UNITED STATES served in these taxa and to comment on their possible swimming behavior.