A Mechanochemical Model of Striae Distensae ⇑ Stephen J
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

UC Davis Dermatology Online Journal
UC Davis Dermatology Online Journal Title Penicillamine-associated cutis laxa and milia en plaque - case report and review of cutaneous changes associated with penicillamine Permalink https://escholarship.org/uc/item/47p4d8zv Journal Dermatology Online Journal, 22(5) Authors Vajdi, Tina Lee, Wiggin Wu Paravar, Taraneh Publication Date 2016 DOI 10.5070/D3225030951 License https://creativecommons.org/licenses/by-nc-nd/4.0/ 4.0 Peer reviewed eScholarship.org Powered by the California Digital Library University of California Volume 22 Number 5 May 2016 Photo Vignette Penicillamine-associated cutis laxa and milia en plaque - case report and review of cutaneous changes associated with penicillamine Tina Vajdi1, Wiggin Wu Lee2, Taraneh Paravar2 Dermatology Online Journal 22 (5): 12 1University of California, San Diego School of Medicine 2Department of Dermatology, University of California, San Diego Correspondence: Taraneh Paravar, MD Assistant Clinical Professor Department of Dermatology University of California, San Diego 8899 University Center Lane, Suite 350 San Diego, California 92122, USA Tel. (858) 657-8322 E-mail: [email protected] Abstract Penicillamine-induced skin changes are rare and include: hypersensitivity reactions, autoimmune reactions, and cutaneous elastoses. We report a case of a 73-year-old man with cystinuria taking penicillamine for over 50 years who presented with penicillamine-induced cutis laxa and milia en plaque. A brief review of penicillamine induced skin changes, specifically cutis laxa and milia en plaque, is presented. Key Words: penicillamine, elastic tissue, cystinuria, cutis laxa, milia en plaque Introduction Penicillamine is a chelating agent commonly used to treat cystinuria and Wilson disease. Cystinuria is a genetic disorder in which patients lack the cysteine amino acid transporter. -
Anetoderma Secondary to Mid-Dermal Elastolysis
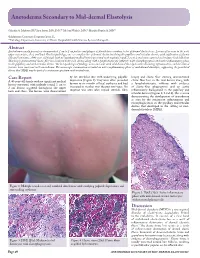
Anetoderma Secondary to Mid-dermal Elastolysis Gabriela A. Maloney, BS,* Jane James, MD, PhD,** Michael Welsch, MD,** Marylee Braniecki, MD** *Midwestern University, Downers Grove, IL **Pathology Department, University of Illinois Hospital & Health Sciences System, Chicago, IL Abstract Anetoderma usually presents as circumscribed, 1 cm to 2 cm patches and plaques of flaccid skin secondary to loss of dermal elastic tissue. Lesions often occur in the neck, upper extremities, chest, and back. On histopathology, one sees complete loss of dermal elastin involving the papillary and reticular dermis, with infiltration of plasma cells and histiocytes. A 40-year-old female with no significant medical history presented with multiple round, 1 cm to 2 cm lesions scattered on her upper back and chest. Skin biopsy demonstrated elastic-fiber loss localized to the mid-dermis along with a lymphohistiocytic infiltrate with elastophagocytosis and active inflammatory phase in the papillary and mid-reticular dermis. The histopathological findings were consistent with mid-dermal elastolysis with advancing inflammation, and the clinical features were consistent with anetoderma. The microscopic examination revealed an active inflammatory phase of mid-dermal elastolysis, supporting the postulated theory that MDE may be part of a continuous spectrum with anetoderma. Case Report by lax, wrinkled skin with underlying palpable biopsy and elastic-fiber staining demonstrated A 40-year-old female with no significant medical depression (Figure 1). They were often preceded elastic-fiber loss in the mid-dermis along with history presented with multiple round, 1 cm to by two to six months of local erythema and had a lymphohistiocytic infiltrate with evidence 2 cm lesions scattered throughout the upper increased in number over the past two years. -

Dermatose Degenerativa Induzida Por D-Penicilamina Em Paciente Com Doença De Wilson
Revista SPDV 76(2) 2018; D-Penicillamine induced degenerative dermopathy; Rui Pedro Santos, Joana Gomes, Celeste Brito. Caso Clínico Dermatose Degenerativa Induzida por D-penicilamina em Paciente com Doença de Wilson Rui Pedro Santos1, Joana Gomes2, Celeste Brito2 1Interno de Dermatovenereologia/Resident, Dermatovenereology, Hospital de Braga, Braga, Portugal 2Especialista de Dermatovenereologia/Specialist of Dermatovenereology, Hospital de Braga, Braga, Portugal RESUMO – As dermatoses degenerativas induzidas por D-penicilamina incluem, entre outras, a elastose perfurante serpiginosa e o pseudo-pseudoxantoma elástico. A elastose perfurante serpiginosa é uma doença perfurante rara caracterizada pela elimi- nação transepidérmica de fibras elásticas anormais. Esta condição pode ser idiopática, reativa ou induzida por D-penicilamina, habitualmente utilizada para o tratamento da doença de Wilson, cistinúria, artrite reumatóide ou esclerose sistémica. Manifesta- ções cutâneas semelhantes a pseudoxantoma elástico mas sem história familiar e mutações do gene ABCC6 foram identificadas como sendo uma dermatose induzida por D-penicilamina e designada de pseudo-pseudoxantoma elástico. Descreve-se o caso de uma mulher de 17 anos tratada por vários anos com D-penicilamina para doença de Wilson, com pápulas assintomáticas, algumas cor de pele e hiperqueratósicas e outras macias e amareladas, na região cervical e face. A histopatolo- gia mostrou a eliminação transepidérmica de fibras elásticas espessadas, em forma de dentes de serra. Estes achados sugeriram uma dermopatia induzida por D-penicilamina e os autores consideraram o diagnóstico de elastose perfurante serpiginosa e pseudo-pseudoxantoma elástico no mesmo paciente. O fármaco foi alterado para acetato de zinco sem lesões novas, mas com manutenção das lesões existentes no seguimento a 1 ano. -

5 Allergic Diseases (And Differential Diagnoses)
Chapter 5 5 Allergic Diseases (and Differential Diagnoses) 5.1 Diseases with Possible IgE Involve- tions (combination of type I and type IVb reac- ment (“Immediate-Type Allergies”) tions). Atopic eczema will be discussed in a separate section (see Sect. 5.5.3). There are many allergic diseases manifesting in The maximal manifestation of IgE-mediated different organs and on the basis of different immediate-type allergic reaction is anaphylax- pathomechanisms (see Sect. 1.3). The most is. In the development of clinical symptoms, common allergies develop via IgE antibodies different organs may be involved and symp- and manifest within minutes to hours after al- toms of well-known allergic diseases of skin lergen contact (“immediate-type reactions”). and mucous membranes [also called “shock Not infrequently, there are biphasic (dual) re- fragments” (Karl Hansen)] may occur accord- action patterns when after a strong immediate ing to the severity (see Sect. 5.1.4). reactioninthecourseof6–12harenewedhy- persensitivity reaction (late-phase reaction, LPR) occurs which is triggered by IgE, but am- 5.1.1 Allergic Rhinitis plified by recruitment of additional cells and 5.1.1.1 Introduction mediators.TheseLPRshavetobedistin- guished from classic delayed-type hypersensi- Apart from being an aesthetic organ, the nose tivity (DTH) reactions (type IV reactions) (see has several very interesting functions (Ta- Sect. 5.5). ble 5.1). It is true that people can live without What may be confusing for the inexperi- breathing through the nose, but disturbance of enced physician is familiar to the allergist: The this function can lead to disease. Here we are same symptoms of immediate-type reactions interested mostly in defense functions against are observed without immune phenomena particles and irritants (physical or chemical) (skin tests or IgE antibodies) being detectable. -

Elastosis Perforans Serpiginosa: a D-Penicillamine Induced Dermatoses in a Patient with Wilson’S Disease
Article / Clinical Case Report Elastosis Perforans Serpiginosa: a D-penicillamine induced dermatoses in a patient with Wilson’s disease Swagatika Samala , Mukund Sablea How to cite: Samal S, Sable M. Elastosis Perforans Serpiginosa: a D-penicillamine induced dermatoses in a patient with Wilson’s disease. Autops Case Rep [Internet]. 2020 Apr-Jun;10(2):e2020167. https://doi.org/10.4322/acr.2020.167 ABSTRACT Long term use of D-penicillamine for Wilson’s disease can be associated with many adverse reactions and systemic side effects. We report the case of a 28-year-old male patient diagnosed with Wilson’s disease presenting with a serpiginous raised violaceous skin lesion in the anterior aspect of the neck over the last six months and two small papules with central umbilication during the last month. Histopathological examination of skin lesions demonstrated transepidermal perforating channel, and the Verhoeff’s-van Gieson stain showed marked increase number of irregular serrated elastic fibers suggesting the diagnosis of D- penicillamine induced elastosis perforans serpiginosa. Keywords Skin Diseases; Biopsy; Elastic tissue. INTRODUCTION CASE REPORT D-penicillamine (DPA) therapy is the mainstay A 28-year-male diagnosed with WD on oral DPA of chelation therapy for patients of Wilson’s therapy (250 mg thrice daily) for the last 18 years disease (WD). Various systemic adverse effects, presented with serpiginous raised violaceous skin including many dermatological manifestations, lesions in the anterior aspect of neck over the last may be observed with prolonged use of this drug. six months and two small papules with central The dermatological side effects of DPA can be of three umbilication for one month (Figure 1). -

Atypical Acrodermatitis Chronica Atrophicans Herxheimer
www.symbiosisonline.org Symbiosis www.symbiosisonlinepublishing.com Case Report Clinical Research in Dermatology: Open Access Open Access Atypical Acrodermatitis Chronica Atrophicans Herxheimer Wollina U1*, Boldt S1, Heinig B2, Schönlebe J3 1Department of Dermatology and Allergology 2Center of Physical and Rehabilitative Medicine 3Institute of Pathology “Georg Schmorl”, Academic Teaching Hospital Dresden-Friedrichstadt, Dresden, Germany Received: December 14, 2015; Accepted: December 19, 2015; Published: December 23, 2015 *Corresponding author: Prof. Dr. U. Wollina, Department of Dermatology and Allergology, Academic Teaching Hospital Dresden-Friedrichstadt, Friedrichstrasse 41, 01067 Dresden, Germany. E-mail: [email protected] Abstract Acrodermatitis Chronica Atrophicans Herxheimer (ACA) is Sensitivity and specificity of enzyme immuno assay and immune a tick-born disease due to infection by Borrelia afzelii, the major blot are 95% and 80-95% for ACA [4]. Polymerase chain reaction vector organism is Ixodes rhicinus. We report on a 48-year-old male (PCR) of skin biopsies was positive in up to 88% on fresh-frozen tissueCase butReport only in 44-52% using paraffin-embedded tissue [5]. symmetric plaques associated with hyperpigmented widely distributedpatient who lesions developed within extensive the tension livid-erythematous lines, and acrocyanosis. fibrosclerotic The of a skin biopsy and laboratory investigations with positive IgG and A 48-year-old male patient was referred to our hospital IgMdiagnosis immunoblots. of ACA has The been patient confirmed was treated by histopathologic by intravenous examination ceftriaxone because of large livid-erythematous fibrosclerotic plaques on resulting in partial remission of cutaneous and extracutaneous his trunk and extremities which developed within half a year. He symptoms. suffered from arterial hypertension and had a penicillin allergy. -

Resident's Page
Resident’s Page SScarscars iinn ddermatology:ermatology: CClinicallinical signisignifi ccanceance BB.. AAnitha,nitha, SS.. RRagunatha,agunatha, AArunrun CC.. IInamadarnamadar Department of Dermatology, Venereology and Leprosy, BLDEA’s SBMP Medical College, Hospital and Research Centre, Bijapur, Karnataka, India AAddressddress fforor ccorrespondenceorrespondence : Dr. Arun C. Inamadar, Professorand Head, Department of Dermatology, Venereology and Leprosy, BLDEA’s SBMP Medical College, Hospital and Research Centre, Bijapur - 586103, Karnataka, India. E-mail:[email protected] [2] A scar is a scar is a scar and only a scar if you don’t ask ß1 protects the collagen from degradation. why” - Shelly and Shelly CCLASSIFICATIONLASSIFICATION OOFF SSCARSCARS[[3]3] A scar is a fibrous tissue replacement that develops as a 1. Fine line scars: Surgical scars consequence of healing at the site of a prior ulcer or 2. Wide (stretched) scars: These develop when fine wound. Cutaneous scarring is a macroscopic disturbance of line surgical scars gradually become stretched the normal structure and function of the skin architecture and widened. They are typically flat, pale, soft, manifesting itself as an elevated or depressed area, with an symptomless scars. Abdominal striae of pregnancy alteration of skin texture, color, vascularity, nerve supply can be considered as variants of these. [1] and biomechanical properties. 3. Atrophic scars: These are flat or depressed below the surrounding skin. They are generally small and Histologically, dermal scars are characterized by thickened often round with an indented or inverted centre. epidermis with a flattened dermo-epidermal junction and They commonly arise after acne or chickenpox. an abnormal organization of the dermal matrix into parallel 4. -

Global Development Assistance for Adolescent Health from 2003 to 2015
Supplementary Online Content Li Z, Li M, Patton GC, Lu C. Global development assistance for adolescent health from 2003 to 2015. JAMA Netw Open. 2018;1(4):e181072. doi:10.1001/jamanetworkopen.2018.1072 eTable 1. List of Donor Countries Included in the CRS eTable 2. 132 Recipients in the CRS (According to the World Health Organization Regions) eTable 3. Key Words to Identify the Related Age Group (Adolescence) in the Creditor Reporting System eTable 4. CRS Purpose Name and Respective Fractions Allocated to Adolescent Health eTable 5. Definitions of DAAH on the Leading Causes of DALYs of Adolescent Health eTable 6. Key Words Used to Search for Projects on Skin and Subcutaneous Diseases in the Creditor Reporting System eTable 7. Key Words Used to Search for Road Injury Projects in the Creditor Reporting System eTable 8. Key Words Used to Search for HIV/AIDS Projects in the Creditor Reporting System eTable 9. Key Words Used to Search for Projects on Iron-Deficiency Anemia in the Creditor Reporting System eTable 10. Key Words Used to Search for Self-Harm Projects in the Creditor Reporting System eTable 11. Key Words Used to Search for Projects on Interpersonal Violence in the Creditor Reporting System eTable 12. Key Words Used to Search for Projects on Depressive Disorders in the Creditor Reporting System eTable 13. Key Words Used to Search for Projects on Lower Back and Neck Pain in the Creditor Reporting System eTable 14. Key Words Used to Search for Diarrheal Projects in the Creditor Reporting System eTable 15. Key Words Used to Search for Tuberculosis Projects in the Creditor Reporting System eTable 16. -

Atrophodermalike Guttate Morphea
CASE LETTER Atrophodermalike Guttate Morphea Nicholas A. Ross, MD; Jason B. Lee, MD; Matthew S. Keller, MD associated arthropod assault. She was treated by an out- PRACTICE POINTS side dermatologist without result for presumed tinea ver- • Atrophodermalike guttate morphea is a potentially sicolor. A follow-up superficial shave biopsy cited subtle underreported or undescribed entity consisting of a psoriasiform dermatitis. Topical steroids did not improve combination of clinicopathologic features. the lesions. Her medical history also was remarkable for a • Widespread hypopigmented macules on the reportedly unprovoked complete rotator cuff tear. trunk and extremities marked by thinned collagen, Physical examination revealed 0.5- to 2.0-cm, ill- fibroplasia, and altered fragmented elastin in the defined, perifollicular and nonfollicular, slightly scaly papillary dermis and upper reticular dermis are the macules and patchescopy on the trunk, arms, and legs. There key features. was no follicular plugging (Figure 1A). The hands, feet, • Atrophoderma, morphea, and lichen sclerosus face, and mucosal surfaces were spared. She had no et atrophicus should be ruled out during clinical workup. family history of similar lesions. Although atrophic in appearance, a single lesion on the left thigh was palpably depressednot (Figure 1B). Serology demonstrated a normal complete blood cell count and comprehensive meta- To the Editor: bolic panel, and negative Lyme titers. Light therapy and Morphea, atrophoderma, guttate lichen sclerosus et atro- topical steroids failed to improve the lesions; calcipotriene phicus (LS&A), anetoderma, and their subtypesDo are cream 0.005% made the lesions erythematous and pruritic. inflammatory processes ultimately leading to dermal A biopsy from a flank lesion demonstrated a normal remodeling. -

Boards' Fodder
boards’ fodder Sound-alikes in dermatology by Jeffrey Kushner, DO, and Kristen Whitney, DO Disease Entity Description Actinic granuloma/ Annular elastolytic Variant of granuloma annulare on sun-damaged skin; annular erythematous giant cell granuloma plaques with slightly atrophic center in sun-exposed areas, which may be precipi- tated by actinic damage. Actinic prurigo PMLE-like disease with photodistributed erythematous papules or nodules and hemorrhagic crust and excoriation. Conjunctivitis and cheilitis are commonly found. Seen more frequently in Native Americans (especially Mestizos). Actinomycetoma “Madura Foot”; suppurative infection due to Nocaria, Actinomadura, or Streptomyces resulting in tissue tumefaction, draining sinuses and extrusion of grains. Actinomycosis “Lumpy Jaw”; Actinomyces israelii; erythematous nodules at the angle of jaw leads to fistulous abscess that drain purulent material with yellow sulfur granules. Acrokeratosis verruciformis Multiple skin-colored, warty papules on the dorsal hands and feet. Often seen in conjunction with Darier disease. Acrodermatitis enteropathica AR; SLC39A4 mutation; eczematous patches on acral, perineal and periorificial skin; diarrhea and alopecia; secondary to zinc malabsorption. Atrophoderma 1) Atrophoderma vermiculatum: Pitted atrophic scars in a honeycomb pattern around follicles on the face; associated with Rombo, Nicolau-Balus, Tuzun and Braun-Falco-Marghescu syndromes. 2) Follicular atrophoderma: Icepick depressions at follicular orifices on dorsal hands/feet or cheeks; associated with Bazex-Dupré-Christol and Conradi- Hünermann-Happle syndromes. 3) Atrophoderma of Pasini and Pierini: Depressed patches on the back with a “cliff-drop” transition from normal skin. 4) Atrophoderma of Moulin: Similar to Pasini/Pierini, except lesions follow the lines of Blaschko. Anetoderma Localized area of flaccid skin due to decreased or absent elastic fibers; exhibits “buttonhole” sign. -

What Is Your Diagnosis?
PHOTO QUIZ What Is Your Diagnosis? A 14-year-old adolescent boy presented with a 2-year history of asymptomatic, soft, outpocketing papules on his abdomen. The lesions disappeared when in a supine position. PLEASE TURN TO PAGE 215 FOR DISCUSSION Leslie E. Stapp, DO, Department of Flight Medicine, Misawa Air Force Base, Japan. The author reports no conflict of interest. 208 CUTIS® Photo Quiz Discussion The Diagnosis: Primary Anetoderma he patient presented to the clinic with or immunologic disorders. Results of the patient’s unusual skin bumps (Figures 1 and 2). Prior laboratory workup were negative or within refer- T to their appearance, he denied any other skin ence range, specifically for human immunode- abnormalities, such as inflammation, erythema, or ficiency virus, rapid plasma reagin, cardiolipin urticaria, on his abdomen. Upon examination, IgG and IgM, C3, C4, antinuclear antibody titer, the patient had average mental development lupus anticoagulant, protein C and S, thyroid- and no physical abnormalities. Skin examination stimulating hormone, and protein electrophoresis. revealed lesions that were soft saclike papules A 4-mm punch biopsy of 2 of the lesions was on the abdomen only, varying from 0.5 to 2 cm performed. The results for both biopsies showed (Figure 3). There were no sensory changes asso- decreased dermal elastic fibers. More specifi- ciated with the lesions. Otherwise, the patient’s cally, elastic stains suggested a decrease in elastic general health was good. He did not have a his- fibers in the mid dermis. Periodic acid-Schiff tory of medication consumption nor did he have stains were negative for fungal pathogens. -

Granulomatous Slack Skin S.J.C
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Erasmus University Digital Repository Clinical and Laboratory Investigations Dermatology 1998;196:382–391 Received: September 5, 1997 Accepted: November 29, 1997 C.W. van Haselen a J. Toonstra a Granulomatous Slack Skin S.J.C. van der Putte b Report of Three Patients with an Updated Review of the Literature J.J.M. van Dongen d C.L.M. van Hees c W.A. van Vloten a ooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooooo Abstract Departments of Purpose: Granulomatous slack skin (GSS) is a rare cutaneous disorder char- a Dermatology and b Pathology, University Hospital of Utrecht, acterized clinically by the evolution of circumscribed erythematous lax skin c Department of Dermatology, masses, especially in the body folds, and histologically by a granulomatous University Hospital of Leiden, and T-cell infiltrate and loss of elastic fibers. GSS is often associated with preceding d Department of Immunology, or subsequent lymphoproliferative malignancies, especially mycosis fungoides Erasmus University Rotterdam, (MF) and Hodgkin’s disease (HD). No effective treatment is known yet. Wheth- The Netherlands er this entity is a benign disorder, a peculiar host reaction to a malignant lym- phoma, a precursor of malignant lymphoma or an indolent cutaneous T-cell lym- phoma (CTCL) in itself is still a matter of debate. Patients and Methods: The results of the patients with GSS from the Netherlands are compared with the cases reported in the world literature. Results: A female patient had had GSS for 8 years without developing a secondary malignancy. In a second female patient with a histologically confirmed diagnosis of MF, GSS developed 18 years later in the axillary and inguinal folds which had previously been affected by plaque- stage MF lesions.