Sea-Level Rise and the Reduction in Pine Forests in the Florida Keys @ Michael S
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Repurposing the East Coast Railway: Florida Keys Extension a Design Study in Sustainable Practices a Terminal Thesis Project by Jacqueline Bayliss
REPURPOSING THE EAST COAST RAILWAY: FLORIDA KEYS EXTENSION A DESIGN STUDY IN SUSTAINABLE PRACTICES A terminal thesis project by Jacqueline Bayliss College of Design Construction and Planning University of Florida Spring 2016 University of Florida Spring 2016 Terminal Thesis Project College of Design Construction & Planning Department of Landscape Architecture A special thanks to Marie Portela Joan Portela Michael Volk Robert Holmes Jen Day Shaw Kay Williams REPURPOSING THE EAST COAST RAILWAY: FLORIDA KEYS EXTENSION A DESIGN STUDY IN SUSTAINABLE PRACTICES A terminal thesis project by Jacqueline Bayliss College of Design Construction and Planning University of Florida Spring 2016 Table of Contents Project Abstract ................................. 6 Introduction ........................................ 7 Problem Statement ............................. 9 History of the East Coast Railway ...... 10 Research Methods .............................. 12 Site Selection ............................... 14 Site Inventory ............................... 16 Site Analysis.................................. 19 Case Study Projects ..................... 26 Limitations ................................... 28 Design Goals and Objectives .................... 29 Design Proposal ............................ 30 Design Conclusions ...................... 40 Appendices ......................................... 43 Works Cited ........................................ 48 Figure 1. The decommissioned East Coast Railroad, shown on the left, runs alongside the Overseas -

Appendix C - Monroe County
2016 Supplemental Summary Statewide Regional Evacuation Study APPENDIX C - MONROE COUNTY This document contains summaries (updated in 2016) of the following chapters of the 2010 Volume 1-11 Technical Data Report: Chapter 1: Regional Demographics Chapter 2: Regional Hazards Analysis Chapter 4: Regional Vulnerability and Population Analysis Funding provided by the Florida Work completed by the Division of Emergency Management South Florida Regional Council STATEWIDE REGIONAL EVACUATION STUDY – SOUTH FLORIDA APPENDIX C – MONROE COUNTY This page intentionally left blank. STATEWIDE REGIONAL EVACUATION STUDY – SOUTH FLORIDA APPENDIX C – MONROE COUNTY TABLE OF CONTENTS APPENDIX C – MONROE COUNTY Page A. Introduction ................................................................................................... 1 B. Small Area Data ............................................................................................. 1 C. Demographic Trends ...................................................................................... 4 D. Census Maps .................................................................................................. 9 E. Hazard Maps .................................................................................................15 F. Critical Facilities Vulnerability Analysis .............................................................23 List of Tables Table 1 Small Area Data ............................................................................................. 1 Table 2 Health Care Facilities Vulnerability -

• the Seven Mile Bridge (Knight Key Bridge HAER FL-2 Moser Channel
The Seven Mile Bridge (Knight Key Bridge HAER FL-2 Moser Channel Bridge Pacet Channel Viaduct) Linking Several Florida Keys Monroe County }-|/ -i c,.^ • Florida '■ L. f'H PHOTOGRAPHS WRITTEN HISTORICAL AND DESCRIPTIVE DATA Historic American Engineering Record National Park Service Department of the Interior Washington D.C. 20240 • THE SEVEN MILE BRIDGE FL-2 MA e^ Ft. A HISTORIC AMERICAN ENGINEERING RECORD THE SEVEN MILE BRIDGE (Knight Key Bridge-Pigeon Key Bridge-Moser Channel Bridge- Pacet Channel Viaduct) Location: Spanning several Florida Keys and many miles of water this bridge is approximately 110 miles from Miami. It begins at Knight Key at the northeast end and terminates at Pacet Key at the southwest end. UTM 487,364E 476.848E 2,732,303N 2,729,606N # Date of Construction 1909-1912 as a railway bridge. Adapted as a concrete vehicular bridge on U.S. I in 1937-1938. Present Owner: Florida Department of Transpor- tation Hayden Burns Building Tallahassee, Florida 32304 Present Use: Since its conversion as a bridge for vehicles it has been in con- tinually heavy use as U.S. I linking Miami with Key West. There is one through draw span riAcis. rLi— z. \r. z.) at Moser Channel, the connecting channel between the Atlantic Ocean and the Gulf of Mexico. It is presently scheduled to be replaced by the State with con- struction already underway in 1980. Significance At the time the Florida East Coast Railway constructed this bridge it was acclaimed as the longest bridge in the world, an engineering marvel. It we.s the most costly of all Flagler's bridges in the Key West Exten- sion. -

Molasses Key Primitive Camp Trip Paddle Information Sheet Description: the Molasses Keys Are Only a Little Over 2 Miles from the South End of the Seven Mile Bridge
S15LK Day Paddles and Overnight Trips - Lower Keys Molasses Key Primitive Camp Trip Paddle Information Sheet Description: The Molasses Keys are only a little over 2 miles from the south end of the Seven Mile Bridge. Take a break at Money Key about halfway there. Look for Bahama Sea Stars on the way out. There is a lot of other sea life to look for like stingrays and sharks. Many bird species inhabit the islands, the rare white-crowned pigeons nest there, and during spring and fall, migrating warblers and other birds can be seen passing thru. Skill Level: Intermediate Distance/Approximate Time: 2.7 Miles Each Way/1.5 Hours each way Launch Site: Little Duck Key - Wayside Park Ramp Special Considerations: As you paddle towards the Molasses Keys, you might be pushed out if the tide is receding or drawn toward the bridges if the tide is rising. There is a section of choppy water between Money and the Molasses Keys. Just stay calm, keep your eyes on the islands and keep paddling! The Molasses Keys are privately owned is- lands that locals and tourists have been visiting for years. Please respect these pristine keys that the owners keep trash free. Do not take anything and do not leave anything, es- pecially trash but also do not post any signs or structures. Hurricane Irma in September 2017 destroyed the islands with major loss of trees and even extensive loss of land (particularly the beach on the little key). Permission is required for camping. Contact Frank & Monica by messaging from the Facebook page, Friends of the Molasses Keys or calling Monica at 305-394-9252. -

Monroe County Stormwater Management Master Plan
Monroe County Monroe County Stormwater Management Master Plan Prepared for Monroe County by Camp Dresser & McKee, Inc. August 2001 file:///F|/GSG/PDF Files/Stormwater/SMMPCover.htm [12/31/2001 3:10:29 PM] Monroe County Stormwater Management Master Plan Acknowledgements Monroe County Commissioners Dixie Spehar (District 1) George Neugent, Mayor (District 2) Charles "Sonny" McCoy (District 3) Nora Williams, Mayor Pro Tem (District 4) Murray Nelson (District 5) Monroe County Staff Tim McGarry, Director, Growth Management Division George Garrett, Director, Marine Resources Department Dave Koppel, Director, Engineering Department Stormwater Technical Advisory Committee Richard Alleman, Planning Department, South Florida WMD Paul Linton, Planning Department, South Florida WMD Murray Miller, Planning Department, South Florida WMD Dave Fernandez, Director of Utilities, City of Key West Roland Flowers, City of Key West Richard Harvey, South Florida Office U.S. Environmental Protection Agency Ann Lazar, Department of Community Affairs Erik Orsak, Environmental Contaminants, U.S. Fish and Wildlife Service Gus Rios, Dept. of Environmental Protection Debbie Peterson, Planning Department, U.S. Army Corps of Engineers Teresa Tinker, Office of Planning and Budgeting, Executive Office of the Governor Eric Livingston, Bureau Chief, Watershed Mgmt, Dept. of Environmental Protection AB i C:\Documents and Settings\mcclellandsi\My Documents\Projects\SIM Projects\Monroe County SMMP\Volume 1 Data & Objectives Report\Task I Report\Acknowledgements.doc Monroe County Stormwater Management Master Plan Stormwater Technical Advisory Committee (continued) Charles Baldwin, Islamorada, Village of Islands Greg Tindle, Islamorada, Village of Islands Zulie Williams, Islamorada, Village of Islands Ricardo Salazar, Department of Transportation Cathy Owen, Dept. of Transportation Bill Botten, Mayor, Key Colony Beach Carlos de Rojas, Regulation Department, South Florida WMD Tony Waterhouse, Regulation Department, South Florida WMD Robert Brock, Everglades National Park, S. -
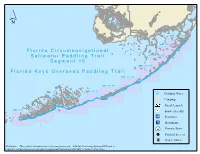
Segment 15 Map Book
´ M M 1 0 4 - 8 9 M M 9 8 - 8 9 Florida Circumnavigational Saltwater Paddling Trail M M 9 1 - 8 1 S e g m e n t 1 5 M M 8 2 - 7 0 Florida Keys Overseas Paddling Trail M M 7 1 - 5 9 M M 6 0 - 4 8 M M 5 0 - 3 9 po Drinking Water M M 2 7 - 1 7 M M 3 9 - 2 7 M M 2 7 - 1 7 t[ Camping M M 1 7 - 7 Kayak Launch M M 8 - 0 M M 8 - 0 Shower Facility I* Restroom I9 Restaurant ²· Grocery Store !e Point of Interest l Hotel / Motel Disclaimer: This guide is intended as an aid to navigation only. A Gobal Positioning System (GPS) unit is required, and persons are encouraged to supplement these maps with NOAA charts or other maps. li d[ li 3 Boggy Key 12 6 Mile Markers 104-96 Bush Point 3 N:25.1252 | W: -80.4054 op [ MM 104 A I* t li Porjoe Key ´ 3 Largo Sound MM 103 6 3 li Whaleback Key A J3 ohn PenneKamp State ParK 6 3 d[ MM 102 li El Radabob Key Swash Keys FL Keys National MM 101 Marine Sanctuary li 6 [ Key Largo d MONROE 6 Shell Key li MM 100 18 d[ 3 Pelican Key MM 99 3 6 li 3 Butternut Key 3 John Pennekamp d[ MM 98 li Coral Reef State Park 6 3 3 MM 97 6 12 6 6 li 6 MM 96 Pigeon Key Verdera Beaclih Rodriguez Key li 0 1 2 4 Miles Sunset Point 12 3 Dove Key li li Mile Markers 98-89 Butternut Key 3 MM 98 A N:25.0242 | W: -80.4943 op I* li 12 3 3 d[ 3 MM 97 ´ 6 li John Pennekamp Bottle Key Coral Reef State Park 12 Stake Key MM 96 6 Pigeon Key li d[ Rodriguez Key 6 Low Key 3 d[ Wild Bird Center MM 95 !e !e li Dove Key MONROd[E 6 12 Tavernier Key MM 94 12 3 li Island Bay Motel !e Dove Creek Conservation Area N: 25.0165 I W: -80.5133 MM 93 3 li d[A Harry -

Outstanding Bridges of Florida*
2013 OOUUTTSSTTAANNDDIINNGG BBRRIIDDGGEESS OOFF FFLLOORRIIDDAA** This photograph collection was compiled by Steven Plotkin, P.E. RReeccoorrdd HHoollddeerrss UUnniiqquuee EExxaammpplleess SSuuppeerriioorr AAeesstthheettiiccss * All bridges in this collection are on the State Highway System or on public roads Record Holders Longest Total Length: Seven Mile Bridge, Florida Keys Second Longest Total Length: Sunshine Skyway Bridge, Lower Tampa Bay Third Longest Total Length: Bryant Patton Bridge, Saint George Island Most Single Bridge Lane Miles: Sunshine Skyway Bridge, Lower Tampa Bay Most Dual Bridge Lane Miles: Henry H. Buckman Bridge, South Jacksonville Longest Viaduct (Bridge over Land): Lee Roy Selmon Crosstown Expressway, Tampa Longest Span: Napoleon Bonaparte Broward Bridge at Dames Point, North Jacksonville Second Longest Span: Sunshine Skyway Bridge, Lower Tampa Bay Longest Girder/Beam Span: St. Elmo W. Acosta Bridge, Jacksonville Longest Cast-In-Place Concrete Segmental Box Girder Span: St. Elmo W. Acosta Bridge, Jacksonville Longest Precast Concrete Segmental Box Girder Span and Largest Precast Concrete Segment: Hathaway Bridge, Panama City Longest Concrete I Girder Span: US-27 at the Caloosahatchee River, Moore Haven Longest Steel Box Girder Span: Regency Bypass Flyover on Arlington Expressway, Jacksonville Longest Steel I Girder Span: New River Bridge, Ft. Lauderdale Longest Moveable Vertical Lift Span: John T. Alsop, Jr. Bridge (Main Street), Jacksonville Longest Movable Bascule Span: 2nd Avenue, Miami SEVEN MILE BRIDGE (new bridge on left and original remaining bridge on right) RECORD: Longest Total Bridge Length (6.79 miles) LOCATION: US-1 from Knights Key to Little Duck Key, Florida Keys SUNSHINE SKYWAY BRIDGE RECORDS: Second Longest Span (1,200 feet), Second Longest Total Bridge Length (4.14 miles), Most Single Bridge Lane Miles (20.7 miles) LOCATION: I–275 over Lower Tampa Bay from St. -

MCTG-2753 ENGLISH Conch Brochure
For More Information KEY LARGO CHAMBER OF COMMERCE (305) The Florida Keys & Key West. Come as you are. FLORIDA KEYS VISITOR CENTER (305) 451-1414 [email protected]/[email protected] Milemarker 106, Overseas Hwy. Key Largo, FL 3451-14143037, U.S. ISLAMORADA CHAMBER OF COMMERCE 305-664-4503305-664-4503/[email protected]@islamoradachamber.org Compose your vacation in any Key. Milemarker 82.6, Overseas Hwy., P.O. Box 915 Islamorada, FL 33036, U.S. MARATHON CHAMBER OF COMMERCE ((305)305 )743-5417 743-5417/[email protected]@floridakeysmarathon.com Milemarker 53, Overseas Hwy. Marathon, FL 33050, U.S. LOWER KEYS CHAMBER OF COMMERCE (305) 872-2411/[email protected] (305) 872-2411 [email protected] KEY LARGO ISLAMORADA MARATHON LOWER KEYS KEY WEST Milemarker 31, Overseas Hwy. Big Pine Key, FL 33040, U.S. Sport Fishing Your Hometown in the KEY WEST CHAMBER OF COMMERCE ((305)305 )294-2587 [email protected]/[email protected] Dive Capital of the World. A Natural Escape. The Uncommon Place. 510 Greene Street, Key West, FL 33040, U.S. Capital of the World. Heart of the Keys. FLORIDA KEYS & KEY WEST TOURIST DEVELOPMENT COUNCIL (305) 296-1552/1-800-FLA-KEYS P.O. Box 866, Key West, FL 33041, U.S. Directions BY AIR: TheThe KeyKey WestWest IInternationalnternationa lAirport Airpor hast ha scommercial commerci aservicel servic ande and is iserveds serve dby b y major airlines. Bmajor airlines.oth the Key We Bothst and the M aKeyrat hWeston a iandrpo rMarathonts have ch airportsarter se rhavevice, chartergenera lservice, aviatio n and rental service. -
Card Sound Water Access Gone by DAVID GOODHUE Access Points Near Card Sheriff Cites Crowds’ Rowdiness, in the Bunch
KEY LARGO 305.451.5700 make. MARATHON 305.743.4397 home. KEY WEST stylish. 305.295.6400 keysfurniture.com WWW.KEYSINFONET.COM SATURDAY, APRIL 12, 2014 VOLUME 61, NO. 30 G 25 CENTS NORTH KEY LARGO Card Sound water access gone By DAVID GOODHUE access points near Card Sheriff cites crowds’ rowdiness, in the bunch. If they had not Monroe County Sheriff’s [email protected] Sound Bridge are now for the come down here and screwed Office spokeswoman Deputy most part off limits. graffiti and trash as the reason it up, nobody would have Becky Herrin called the Card Those wanting to launch a The nearest place for ever cared,” Murphy said. “It Sound area “a hot spot for boat or personal watercraft someone to put his or her Road spots because exces- is also known for drunken gets worse and worse with trouble.” She said deputies from anywhere on the Monroe boat in the water for free in sive weekend parties left the fights, loud music and dan- every passing week.” were frequently called there to County side of Card Sound Monroe County is now about area littered with trash, beer gerous personal-watercraft None of the five small respond to fights, traffic inci- Road must look elsewhere. 40 miles south at the Indian bottles and cans and other drivers. She said the majority bridges that span Card Sound dents and other disturbances. Using Jersey barriers, Key fill ramp at mile marker debris, as well as spray- of people who play there are Road, which are popular fish- Herrin said “Jet Ski boulders and “No Trespass” 79 in Islamorada. -

Motion on the Ocean
MOTION ON THE OCEAN A PUBLIC HEALTH GUIDE TO PHYSICAL ACTIVITY IN PARKS ACROSS THE FLORIDA KEYS #healthiestweightfl Download this booklet on your smartphone at monroe.floridahealth.gov 2 Editorial Board: Executive Lead Team, Florida Department of Health in Monroe County Bob Eadie, Administrator Mark Whiteside MD MPH, Medical Director Mary Vanden Brook, Administrative Services Director Bill Brookman MPH, Community Health Services Director Joan Higgs RN, Nursing Director Mary Turner, ARNP, Nursing Director Map Credits Dr. Derrick Scott and Armando Luzula Department of Global and Socio-Cultural Studies Dr. Joseph Yates Porter Florida International University (1847-1927) Key West Native and Director of Florida’s First State Board of Health Researchers Alison Morales Kerr, MPH, Community Health Program Manager, DOH-Monroe Christopher Tittel, MA, Marketing Director, DOH-Monroe Mark Whiteside, MD, MPH, Medical Director, DOH-Monroe Andrew Bulla, MBA, MS, Administrative Services, DOH-Monroe Courtney Smith, RD, LD, DOH-Monroe Ryan Porterfield, Bachelor of Science in Biotechnology Candidate, University of Florida Kevin Wilson,P.E., Division Director, Public Works & Engineering, Monroe County Roman Gastesi, Administrator, Monroe County Debbie Frederick, CPM, Deputy Administrator, Monroe County Virginia Burgohy Irving, Ed.D., Douglass School Black Educators’ Memorial Project, Inc. Photo Credits Eric Whiteside Amy Grimm Christopher Tittel Danielle Carson Designer Christopher Tittel 3 FROM THE ADMINISTRATOR Welcome to Motion on the Ocean, a guide to parks across the Florida Keys that has been written, photographed, and designed with fitness in mind. In 2013, Dr. John H. Armstrong, State Surgeon General and Secretary of Health, initiated the Healthiest Weight Florida campaign. With an obesity epidemic nationwide, Dr. -

Florida Keys P E!
FLORIDA KEYS AQUEDUCT AUTHORITY BOARD OF DIRECTORS’ MEETING Open to the Public Florida Keys Mosquito Control District 503 107th Street Marathon, Florida 33050 Wednesday, July 26, 2017 10:30 a.m. PUBLIC HEARINGS Presentation on Proposed Budget for Fiscal Year 2018 and Proposed Water Rate Increase REGULAR MEETING AGENDA INVOCATION PLEDGE OF ALLEGIANCE EMPLOYEE AWARDS Juan Lopez – 15 Years of Service Aaron Sipe – 10 Years of Service EMPLOYEE OF THE YEAR Natalie Allen RETIREMENT Mark Theobald APPROVAL OF MINUTES June 28, 2017 Board Meeting ADDITIONS/DELETIONS TO AGENDA ACCEPTANCE OF AGENDA Note: The Board of Directors does allow time for comments from the public, who have completed sign-in cards, on any Agenda Item or any specific areas of concern. Individuals will be allotted 3 minutes to make presentation. Representatives from established groups will be allotted 5 minutes to make presentation. FKAA Agenda – July 26, 2017 Page 2 CONSENT AGENDA ITEMS OFFICE OF COUNSEL 1. OOC – 01 Consideration to approve payment to Bryant, Miller & Olive for professional services rendered and expenses incurred for the month of May 2017 regarding defense of declaratory judgment, Delaney & Kulikowsky vs. Florida Keys Aqueduct Authority and Monroe County ($1,228.06 Monroe County) DIVISION OF UTILITY OPERATIONS 2. DUO – 01 Consideration to approve the abandonment of right-of-way described as a portion of Severino Drive in Islamorada, Florida as shown on Venetian Shores Plat No. 3 according to Plat thereof as recorded in Plat Book 5, Page 48 of the Public Records -

Archaeological and Biological Examination of the Brick Wreck (8MO1881) Off Vaca Key, Monroe County, Florida
Archaeological and Biological Examination of the Brick Wreck (8MO1881) off Vaca Key, Monroe County, Florida Roger C. Smith, Jeffrey T. Moates, Debra G. Shefi, and Brian J. Adams, with Contributions by Brenda S. Altmeier, Lee A. Newsom, and Colleen L. Reese A Report Submitted to the Florida Keys National Marine Sanctuary in Fulfillment of a NOAA Maritime Heritage Program Mini-grant October 2006 Acknowledgments Principal participants included Dr. Roger Smith, Jeffrey Moates, and Debra Shefi of the Florida Bureau of Archaeological Research; Brian Adams of the University of West Florida; Brenda Altmeier, John Halas, Cheva Heck, and Amy Massey of the Florida Keys National Marine Sanctuary; Bruce Terrell, Senior Archaeologist of the NOAA National Marine Sanctuaries Program; and Dr. John Broadwater, Director of the NOAA Maritime Heritage Program. Other participants included KC Smith of the Museum of Florida History; Harry DeLashmutt, Ports Manager, and Richard Tanner, Dockmaster of the City of Marathon Marina; George Garrett, Director of the Monroe County Marine Resources Department; and Bruce Reyngoudt, David Becker, and Sean Meehan of the Florida Keys National Marine Sanctuary. Figure 1. Brick Wreck team (from left to right): Brenda Altmeier, Richard Tanner, Brian Adams, Jeff Moates, Roger Smith, Debra Shefi ii Table of Contents Introduction......................................................................................................................... 1 Research Design and Proposal...................................................................................