Chapter 11A Structuring Herbivore Communities
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Population, Distribution and Conservation Status of Sitatunga (Tragelaphus Spekei) (Sclater) in Selected Wetlands in Uganda
POPULATION, DISTRIBUTION AND CONSERVATION STATUS OF SITATUNGA (TRAGELAPHUS SPEKEI) (SCLATER) IN SELECTED WETLANDS IN UGANDA Biological -Life history Biological -Ecologicl… Protection -Regulation of… 5 Biological -Dispersal Protection -Effectiveness… 4 Biological -Human tolerance Protection -proportion… 3 Status -National Distribtuion Incentive - habitat… 2 Status -National Abundance Incentive - species… 1 Status -National… Incentive - Effect of harvest 0 Status -National… Monitoring - confidence in… Status -National Major… Monitoring - methods used… Harvest Management -… Control -Confidence in… Harvest Management -… Control - Open access… Harvest Management -… Control of Harvest-in… Harvest Management -Aim… Control of Harvest-in… Harvest Management -… Control of Harvest-in… Tragelaphus spekii (sitatunga) NonSubmitted Detrimental to Findings (NDF) Research and Monitoring Unit Uganda Wildlife Authority (UWA) Plot 7 Kira Road Kamwokya, P.O. Box 3530 Kampala Uganda Email/Web - [email protected]/ www.ugandawildlife.org Prepared By Dr. Edward Andama (PhD) Lead consultant Busitema University, P. O. Box 236, Tororo Uganda Telephone: 0772464279 or 0704281806 E-mail: [email protected] [email protected], [email protected] Final Report i January 2019 Contents ACRONYMS, ABBREVIATIONS, AND GLOSSARY .......................................................... vii EXECUTIVE SUMMARY ....................................................................................................... viii 1.1Background ........................................................................................................................... -

Download Publication
Caesar Kleberg A Publication of the Caesar Kleberg Wildlife ResearchTracks Institute CAESAR KLEBERG WILDLIFE RESEARCH INSTITUTE TEXAS A&M UNIVERSITY - KINGSVILLE 1 Caesar Kleberg Volume 5 | Issue 2 | Fall 2020 In This Issue 3 From the Director Tracks 4 Restoration: Just What Do You Mean By That? 8 The Unique Nature of Colonial-nesting Waterbirds 12 Texas Horned Lizard 8 Detectives Learn More About CKWRI The Caesar Kleberg Wildlife Research Institute at 16 Donor Spotlight: Texas A&M University-Kingsville is a Master’s and Mike Reynolds Ph.D. Program and is the leading wildlife research organization in Texas and one of the finest in the nation. Established in 1981 by a grant from the Caesar Kleberg 20 Fawn Survival and Foundation for Wildlife Conservation, its mission is Recruitment in South Texas to provide science-based information for enhancing the conservation and management of Texas wildlife. 23 Alumni Spotlight: J. Dale James Visit our Website www.ckwri.tamuk.edu Caesar Kleberg Wildlife Research Institute Texas A&M University-Kingsville 700 University Blvd., MSC 218 Kingsville, Texas 78363 4 (361) 593-3922 Follow us! Facebook: @CKWRI Twitter: @CKWRI Instagram: ckwri_official Cover Photo by iStock 2 Magazine Design and Layout by Gina Cavazos Caesar Kleberg From the Director The overarching mission of the Caesar Kleberg Wildlife Research Institute is to promote wildlife conservation. We do this primarily by conducting research and producing knowledge to benefit wildlife managers and land stewards. We also promote wildlife conservation by training the next generation of wildlife biologists Tracks through our graduate education programs. However, producing knowledge and trained professionals is not enough. -

Animals of Africa
Silver 49 Bronze 26 Gold 59 Copper 17 Animals of Africa _______________________________________________Diamond 80 PYGMY ANTELOPES Klipspringer Common oribi Haggard oribi Gold 59 Bronze 26 Silver 49 Copper 17 Bronze 26 Silver 49 Gold 61 Copper 17 Diamond 80 Diamond 80 Steenbok 1 234 5 _______________________________________________ _______________________________________________ Cape grysbok BIG CATS LECHWE, KOB, PUKU Sharpe grysbok African lion 1 2 2 2 Common lechwe Livingstone suni African leopard***** Kafue Flats lechwe East African suni African cheetah***** _______________________________________________ Red lechwe Royal antelope SMALL CATS & AFRICAN CIVET Black lechwe Bates pygmy antelope Serval Nile lechwe 1 1 2 2 4 _______________________________________________ Caracal 2 White-eared kob DIK-DIKS African wild cat Uganda kob Salt dik-dik African golden cat CentralAfrican kob Harar dik-dik 1 2 2 African civet _______________________________________________ Western kob (Buffon) Guenther dik-dik HYENAS Puku Kirk dik-dik Spotted hyena 1 1 1 _______________________________________________ Damara dik-dik REEDBUCKS & RHEBOK Brown hyena Phillips dik-dik Common reedbuck _______________________________________________ _______________________________________________African striped hyena Eastern bohor reedbuck BUSH DUIKERS THICK-SKINNED GAME Abyssinian bohor reedbuck Southern bush duiker _______________________________________________African elephant 1 1 1 Sudan bohor reedbuck Angolan bush duiker (closed) 1 122 2 Black rhinoceros** *** Nigerian -

Common Eland
Tragelaphus oryx – Common Eland recognised, though their validity has been in dispute (Thouless 2013): Tragelaphus o. livingstonii (Sclater 1864; Livingstone's Eland): also called kaufmanni, niediecki, selousi and triangularis. It is found in the Central Zambezian Miombo woodlands i.e. south- central Africa (Angola, Zambia, Democratic Republic of the Congo, Zimbabwe, Mozambique and Malawi). Livingstone's Eland has a brown pelt with up to twelve stripes. Tragelaphus o. oryx (Pallas 1766; Cape Eland): also called alces, barbatus, canna and oreas. This subspecies is found south of the Zambezi river (South Africa, Botswana and Namibia). The fur is tawny, and adults lose their stripes. Regional Red List status (2016) Least Concern Tragelaphus o. pattersonianus (Lydekker 1906; East National Red List status (2004) Least Concern African Eland or Patterson's Eland): also called Reasons for change No change billingae. It is found in east Africa extending into the Somali arid areas, hence its common name. Its coat Global Red List status (2008) Least concern can have up to 12 stripes. TOPS listing (NEMBA) None Tragelaphus o. oryx occurs throughout the larger part of South Africa, but the far northern Limpopo Province CITES listing None bordering Zimbabwe is regarded as a transitional zone Endemic No between T. o. oryx and T. o. livingstonii or an area where they overlap. This argues the case that they should rather During drought conditions Eland roam extensively be described as ecotypes (in ecotypes, it is common for in order to meet forage and water requirements; in continuous, gradual geographic variation to impose the southern Kalahari during abnormally dry analogous phenotypic and/or genetic variation; this conditions, Eland were found to cover more than situation is called cline.). -

Mammal Species Richness at a Catena and Nearby Waterholes During a Drought, Kruger National Park, South Africa
diversity Article Mammal Species Richness at a Catena and Nearby Waterholes during a Drought, Kruger National Park, South Africa Beanélri B. Janecke Animal, Wildlife & Grassland Sciences, University of the Free State, 205 Nelson Mandela Road, Park West, Bloemfontein 9301, South Africa; [email protected]; Tel.: +27-51-401-9030 Abstract: Catenas are undulating hillslopes on a granite geology characterised by different soil types that create an environmental gradient from crest to bottom. The main aim was to determine mammal species (>mongoose) present on one catenal slope and its waterholes and group them by feeding guild and body size. Species richness was highest at waterholes (21 species), followed by midslope (19) and sodic patch (16) on the catena. Small differences observed in species presence between zones and waterholes and between survey periods were not significant (p = 0.5267 and p = 0.9139). In total, 33 species were observed with camera traps: 18 herbivore species, 10 carnivores, two insectivores and three omnivores. Eight small mammal species, two dwarf antelopes, 11 medium, six large and six mega-sized mammals were observed. Some species might not have been recorded because of drought, seasonal movement or because they travelled outside the view of cameras. Mammal presence is determined by food availability and accessibility, space, competition, distance to water, habitat preferences, predators, body size, social behaviour, bound to territories, etc. The variety in body size and feeding guilds possibly indicates a functioning catenal ecosystem. This knowledge can be beneficial in monitoring and conservation of species in the park. Keywords: catena ecosystem; ephemeral mud wallows; habitat use; mammal variety; Skukuza area; Citation: Janecke, B.B. -

History of Insular Ecology and Biogeography - Harold Heatwole
OCEANS AND AQUATIC ECOSYSTEMS- Vol. II - History of Insular Ecology and Biogeography - Harold Heatwole HISTORY OF INSULAR ECOLOGY AND BIOGEOGRAPHY Harold Heatwole North Carolina State University, Raleigh, North Carolina, USA Keywords: islands; insular dynamics; One Tree Island; Aristotle; Darwin; Wallace; equilibrium theory; biogeography; immigration; extinction; species-turnover; stochasticism; determinism; null hypotheses; trophic structure; transfer organisms; assembly rules; energetics. Contents 1. Ancient and Medieval Concepts: the Birth of Insular Biogeography 2. Darwin and Wallace: the Dawn of the Modern Era 3. Genetics and Insular Biogeography 4. MacArthur and Wilson: the Equilibrium Theory of Insular Biogeography 5. Documenting and Testing the Equilibrium Theory 6. Modifying the Equilibrium Theory 7. Determinism versus Stochasticism in Insular Communities 8. Insular Energetics and Trophic Structure Stability Glossary Bibliography Biographical Sketch Summary In ancient times, belief in spontaneous generation and divine creation dominated thinking about insular biotas. Weaknesses in these theories let to questioning of those ideas, first by clergy and later by scientists, culminating in Darwin's and Wallace's postulation of evolution through natural selection. MacArthur and Wilson presented a dynamic equilibrium model of insular ecology and biogeography that represented the number of species on an island as an equilibrium between immigration and extinction rates, as influenced by insular sizes and distances from mainlands. This model has been tested and found generally true but requiring minor modifications, such as accounting for disturbancesUNESCO affecting the equilibrium nu–mber. EOLSS There has been basic controversy as to whether insular biotas reflect deterministic or stochastic processes. It is likely that neither extreme SAMPLEis completely correct but rather CHAPTERS the two kinds of processes interact. -

Brochure Highlight Those Impressive Russia
2019 44 years and counting The products and services listed Join us on Facebook, follow us on Instagram or visit our web site to become one Table of Contents in advertisements are offered and of our growing number of friends who receive regular email updates on conditions Alaska . 4 provided solely by the advertiser. and special big game hunt bargains. Australia . 38 www.facebook.com/NealAndBrownleeLLC Neal and Brownlee, L.L.C. offers Austria . 35 Instagram: @NealAndBrownleeLLC no guarantees, warranties or Azerbaijan . 31 recommendations for the services or Benin . 18 products offered. If you have questions Cameroon . 19 related to these services, please contact Canada . 6 the advertiser. Congo . 20 All prices, terms and conditions Continental U .S . 12 are, to the best of our knowledge at the Ethiopia . 20 time of printing, the most recent and Fishing Alaska . 42 accurate. Prices, terms and conditions Fishing British Columbia . 41 are subject to change without notice Fishing New Zealand . 42 due to circumstances beyond our Kyrgyzstan . 31 control. Jeff C. Neal Greg Brownlee Trey Sperring Mexico . 14 Adventure travel and big game 2018 was another fantastic year for our company thanks to the outfitters we epresentr and the Mongolia . 32 hunting contain inherent risks and clients who trusted us. We saw more clients traveling last season than in any season in the past, Mozambique . 21 dangers by their very nature that with outstanding results across the globe. African hunting remained strong, with our primary Namibia . 22 are beyond the control of Neal and areas producing outstanding success across several countries. Asian hunting has continued to be Nepal . -

Mixed-Species Exhibits with Pigs (Suidae)
Mixed-species exhibits with Pigs (Suidae) Written by KRISZTIÁN SVÁBIK Team Leader, Toni’s Zoo, Rothenburg, Luzern, Switzerland Email: [email protected] 9th May 2021 Cover photo © Krisztián Svábik Mixed-species exhibits with Pigs (Suidae) 1 CONTENTS INTRODUCTION ........................................................................................................... 3 Use of space and enclosure furnishings ................................................................... 3 Feeding ..................................................................................................................... 3 Breeding ................................................................................................................... 4 Choice of species and individuals ............................................................................ 4 List of mixed-species exhibits involving Suids ........................................................ 5 LIST OF SPECIES COMBINATIONS – SUIDAE .......................................................... 6 Sulawesi Babirusa, Babyrousa celebensis ...............................................................7 Common Warthog, Phacochoerus africanus ......................................................... 8 Giant Forest Hog, Hylochoerus meinertzhageni ..................................................10 Bushpig, Potamochoerus larvatus ........................................................................ 11 Red River Hog, Potamochoerus porcus ............................................................... -
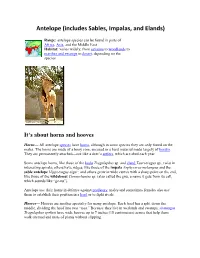
Antelope (Includes Sables, Impalas, and Elands)
Antelope (includes Sables, Impalas, and Elands) Range: antelope species can be found in parts of Africa, Asia, and the Middle East Habitat: varies widely, from savanna to woodlands to marshes and swamps to desert, depending on the species It’s about horns and hooves Horns— All antelope species have horns, although in some species they are only found on the males. The horns are made of a bony core, encased in a hard material made largely of keratin. They are permanently attached—not like a deer’s antlers, which are shed each year. Some antelope horns, like those of the kudu Tragelaphus sp. and eland Taurotragus sp., twist in interesting spirals; others have ridges, like those of the impala Aephyceros melampus and the sable antelope Hippotragus niger; and others grow in wide curves with a sharp point on the end, like those of the wildebeest Connochaetes sp. (also called the gnu, a name it gets from its call, which sounds like “ge-nu”). Antelope use their horns in defense against predators; males and sometimes females also use them to establish their position in a herd or to fight rivals Hooves— Hooves are another specialty for many antelope. Each hoof has a split down the middle, dividing the hoof into two “toes.” Because they live in wetlands and swamps, sitatungas Tragelaphus spekeii have wide hooves up to 7 inches (18 centimeters) across that help them walk on mud and mats of plants without slipping. Nile lechwes Kobus magaceros, which also live in swampy areas, have long, pointed hooves to give them sure footing in the water. -

Society for Ecological Restoration Intern a T I O N
society for ecological restoration intern a t i o n a l The SER International Primer o n Ecological Restoration Society for Ecological Restoration International Science & Policy Working Group (Version 2: October, 2004)* Section 1: Overview . 1 Section 2: Definition of Ecological Restoration . 3 Section 3: Attributes of Restored Ecosystems . 3 Section 4: Explanations of Terms . 4 Section 5: Reference Ecosystems . 8 Section 6: Exotic Species . 9 Section 7: Monitoring and Evaluation . 10 Section 8: Restoration Planning. 11 Section 9: Relationship Between Restoration Practice and Restoration Ecology . 11 Section 10: Relationship of Restoration to Other Activities . 12 Section 11: Integration of Ecological Restoration into a Larger Program. 13 This document should be cited as: Society for Ecological Restoration International Science & Policy Working Group. 2004. The SER International Primer on Ecological Restoration. www.ser.org & Tucson: Society for Ecological Restoration International. The principal authors of this Primer were André Clewell (Quincy, FL USA), James Aronson (Montpellier, France), and Keith Winterhalder (Sudbury, ON Canada). Clewell initially proposed the Primer and wrote its first draft. Aronson and Winterhalder, in collaboration with Clewell, revised the Primer into its present form. Winterhalder, in his capacity as Chairperson of SER’s Science & Policy Working Group, coordinated this effort and invited other Working Group members to participate. Eric Higgs (Victoria, BC Canada) crafted the Overview sec- tion. Dennis Martinez (Douglas City, CA USA) contributed a position paper that became the basis for text pertaining to cultural ecosystems. Other Working Group members provided critiques and suggestions as the work progressed, including Richard Hobbs (Murdoch, WA Australia), James Harris (London, UK), Carolina Murcia (Cali, Colombia), and John Rieger (San Diego, CA USA). -

Assembly Rules for Functional Groups of North American Shrews: Effects of Geographic Range and Habitat Partitioning
OIKOS 107: 141Á/147, 2004 Assembly rules for functional groups of North American shrews: effects of geographic range and habitat partitioning Timothy S. McCay, Matthew J. Lovallo, W. Mark Ford and Michael A. Menzel McCay, T. S., Lovallo, M. J., Ford, W. M. and Menzel, M. A. 2004. Assembly rules for functional groups of North American shrews: effects of geographic range and habitat partitioning. Á/ Oikos 107: 141Á/147. We examined the representation of shrew species within assemblages at 197 sites in the southern Appalachian Mountains, USA. Assemblages were classified according to representation of functional groups, including fossorial, small epigeal, and large epigeal. Average (9/SD) species richness was 2.99/1.0 and assemblages averaged 0.8 species in the fossorial and large epigeal groups and 1.3 species in the small epigeal group. Compliance with Fox’s assembly rule was evaluated by dividing sites into those likely under the rule (favored) and those unlikely under the rule. The number of favored sites was compared to expected numbers of favored sites under three null models of community development, which variously incorporated observed characteristics of species in the regional pool. Number of favored sites (175 [89%]) exceeded the number found in each of 25,000 sets of assemblages simulated (PB/0.00004) using algorithms that incorporated the observed species richness of each site and the observed incidences of each species. However, the number of favored sites did not differ (P/0.1054) from the number expected when patterns of allopatry and sympatry were preserved in the null model. Thus, the tendency for the equitable representation of functional groups in shrew assemblages was primarily due to the pattern of allopatry among similar species, and we believe that the striking compliance of these sites with Fox’s assembly rule is largely due to structure within the regional species pool, rather than extant competitive interactions. -

A REVIEW on ECOLOGICAL ASSEMBLY RULES Revista EIA, Vol
Revista EIA ISSN: 1794-1237 [email protected] Escuela de Ingeniería de Antioquia Colombia Colorado Zuluaga, Gabriel Jaime HOW ECOLOGICAL COMMUNITIES ARE STRUCTURED: A REVIEW ON ECOLOGICAL ASSEMBLY RULES Revista EIA, vol. 12, núm. 24, julio-diciembre, 2015, pp. 27-53 Escuela de Ingeniería de Antioquia Envigado, Colombia Available in: http://www.redalyc.org/articulo.oa?id=149244222002 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative Revista EIA, ISSN 1794-1237 Revista impresa - ISSN WEB 2463-0950 / Año XII / Volumen 12 / Número 24 / Julio-diciembre 2015 / pp. 27-53 / Publicación semestral de carácter técnico-científico / Escuela de Ingeniería de Antioquia —EIA—, Envigado (Colombia) HOW ECOLOGICAL COMMUNITIES ARE STRUCTURED: A REVIEW ON ECOLOGICAL ASSEMBLY RULES 1 ABSTRACT Gabriel Jaime Colorado Zuluaga Whether biological communities are deterministic or stochastic assemblages of species has long been a central topic of ecology. The widely demonstrated presence of structural patterns in nature may imply the existence of rules that regulate the organization of ecological communities. In this review, I present a compilation of major assembly rules that fundament, in a great proportion, the community assembly theory. Initially, I present a general overview of key concepts underlying mechanisms in the structure of ecological communities. Subsequently, two major approaches or paradigms associated to the assembly of communities, in particular the origin of assembly rules, definition, the problem of scale and (i.e. species-based and trait-based) for the assembly of communities are discussed.