Novel Transcriptional Networks Regulated by CLOCK in Human Neurons
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Basic Helix-Loop-Helix Transcription Factors DEC1 and DEC2 Regulate the Paclitaxel-Induced Apoptotic Pathway of MCF-7 Human Breast Cancer Cells
INTERNATIONAL JOURNAL OF MoleCular MEDICine 27: 491-495, 2011 Basic helix-loop-helix transcription factors DEC1 and DEC2 regulate the paclitaxel-induced apoptotic pathway of MCF-7 human breast cancer cells YUNYAN WU1,2, FUYUKI SATO1, UJJAL KUMAR BHAWAL3, TAKESHI KAWAMOTO4, KATSUMI FUJIMOTO4, MITSUHIDE NOSHIRO4, SATOKO MOROHASHI1, YUKIO KATO4 and HIROSHI KIJIMA1 1Department of Pathology and Bioscience, Hirosaki University Graduate School of Medicine, Hirosaki 036-8562, Japan; 2Department of Pathology, College of Basic Medical Sciences, China Medical University, Shenyang 110001, P.R. China; 3Department of Oral and Maxillofacial Surgery and High-Tech Research Center, Kanagawa Dental College, Yokosuka 238-8580; 4Department of Dental and Medical Biochemistry, Hiroshima University Graduate School of Biomedical Science, Hiroshima 734-8553, Japan Received November 17, 2010; Accepted January 10, 2011 DOI: 10.3892/ijmm.2011.617 Abstract. Differentiated embryonic chondrocyte gene Introduction (DEC) 1 (BHLHE40/Stra13/Sharp2) and DEC2 (BHLHE41/ Sharp1) are basic helix-loop-helix (bHLH) transcription Paclitaxel is an anti-tumor drug that is used against a wide factors that are associated with the regulation of apoptosis, cell variety of solid tumors (1,2) and affects the expression of proliferation and circadian rhythms, as well as malignancy Bcl-2, Bcl-xL and phosphorylated-c-Jun NH2-terminal kinase in various cancers. However, the roles of DEC1 and DEC2 (JNK), and the activation of caspases and poly (ADP-ribose) expression in breast cancer are poorly understood. In this polymerase PARP (3-6), resulting in the induction of apop- study, we sought to examine the roles of DEC1 and DEC2 tosis by p53-dependent or -independent mechanisms (7-9). -

Downregulation of Dipeptidyl Peptidase 4 Accelerates Progression to Castration-Resistant Prostate Cancer Joshua W
Published OnlineFirst September 21, 2018; DOI: 10.1158/0008-5472.CAN-18-0687 Cancer Priority Report Research Downregulation of Dipeptidyl Peptidase 4 Accelerates Progression to Castration-Resistant Prostate Cancer Joshua W. Russo1,CeGao2, Swati S. Bhasin2, Olga S. Voznesensky1, Carla Calagua3, Seiji Arai1,4, Peter S. Nelson5, Bruce Montgomery6, Elahe A. Mostaghel5, Eva Corey6, Mary-Ellen Taplin7, Huihui Ye3, Manoj Bhasin2, and Steven P. Balk1 Abstract The standard treatment for metastatic prostate cancer, cleaves dipeptides from multiple growth factors, resulting in androgen deprivation therapy (ADT), is designed to suppress their increased degradation. DPP4 mRNA and protein were androgen receptor (AR) activity. However, men invariably also decreased in clinical CRPC cases, and inhibition of progress to castration-resistant prostate cancer (CRPC), and DPP4 with sitagliptin enhanced the growth of prostate AR reactivation contributes to progression in most cases. To cancer xenografts following castration. Significantly, DPP4 identify mechanisms that may drive CRPC, we examined a inhibitors are frequently used to treat type 2 diabetes as they VCaP prostate cancer xenograft model as tumors progressed increase insulin secretion. Together, these results implicate from initial androgen sensitivity prior to castration to castra- DPP4 as an AR-regulated tumor suppressor gene whose tion resistance and then on to relapse after combined therapy loss enhances growth factor activity and suggest that treat- with further AR-targeted drugs (abiraterone plus enzaluta- ment with DPP4 inhibitors may accelerate emergence of mide). AR activity persisted in castration-resistant and abir- resistance to ADT. aterone/enzalutamide–resistant xenografts and was associated with increased expression of the AR gene and the AR-V7 splice Significance: These findings identify DPP4 as an AR-stim- variant. -

The Title of the Dissertation
UNIVERSITY OF CALIFORNIA SAN DIEGO Novel network-based integrated analyses of multi-omics data reveal new insights into CD8+ T cell differentiation and mouse embryogenesis A dissertation submitted in partial satisfaction of the requirements for the degree Doctor of Philosophy in Bioinformatics and Systems Biology by Kai Zhang Committee in charge: Professor Wei Wang, Chair Professor Pavel Arkadjevich Pevzner, Co-Chair Professor Vineet Bafna Professor Cornelis Murre Professor Bing Ren 2018 Copyright Kai Zhang, 2018 All rights reserved. The dissertation of Kai Zhang is approved, and it is accept- able in quality and form for publication on microfilm and electronically: Co-Chair Chair University of California San Diego 2018 iii EPIGRAPH The only true wisdom is in knowing you know nothing. —Socrates iv TABLE OF CONTENTS Signature Page ....................................... iii Epigraph ........................................... iv Table of Contents ...................................... v List of Figures ........................................ viii List of Tables ........................................ ix Acknowledgements ..................................... x Vita ............................................. xi Abstract of the Dissertation ................................. xii Chapter 1 General introduction ............................ 1 1.1 The applications of graph theory in bioinformatics ......... 1 1.2 Leveraging graphs to conduct integrated analyses .......... 4 1.3 References .............................. 6 Chapter 2 Systematic -

A Common Analgesic Enhances the Anti-Tumour Activity of 5-Aza-2’- Deoxycytidine Through Induction of Oxidative Stress
bioRxiv preprint doi: https://doi.org/10.1101/2020.03.31.017947; this version posted April 1, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. A common analgesic enhances the anti-tumour activity of 5-aza-2’- deoxycytidine through induction of oxidative stress Hannah J. Gleneadie1,10, Amy H. Baker1, Nikolaos Batis2, Jennifer Bryant2, Yao Jiang3, Samuel J.H. Clokie4, Hisham Mehanna2, Paloma Garcia5, Deena M.A. Gendoo6, Sally Roberts5, Alfredo A. Molinolo7, J. Silvio Gutkind8, Ben A. Scheven1, Paul R. Cooper1, Farhat L. Khanim9 and Malgorzata Wiench1, 5,*. 1School of Dentistry, Institute of Clinical Studies, College of Medical and Dental Sciences, The University of Birmingham, Birmingham, B5 7EG, UK; 2Institute of Head and Neck Studies and Education (InHANSE), The University of Birmingham, Birmingham, B15 2TT, UK; 3School of Biosciences, The University of Birmingham, Birmingham, B15 2TT, UK; 4West Midlands Regional Genetics Laboratory, Birmingham Women’s and Children’s Hospital, Birmingham, B15 2TG, UK; 5Institute of Cancer and Genomic Sciences, College of Medical and Dental Sciences, The University of Birmingham, Birmingham, B15 2TT, UK; 6Centre for Computational Biology, Institute of Cancer and Genomic Sciences, The University of Birmingham, Birmingham, B15 2TT, UK; 7Moores Cancer Center and Department of Pathology, University of California San Diego, La Jolla, CA 92093, USA; 8Department of Pharmacology and Moores Cancer -

Effects of Circadian Clock Genes and Health-Related
RESEARCH ARTICLE Effects of circadian clock genes and health- related behavior on metabolic syndrome in a Taiwanese population: Evidence from association and interaction analysis Eugene Lin1,2,3*, Po-Hsiu Kuo4, Yu-Li Liu5, Albert C. Yang6,7, Chung-Feng Kao8, Shih- Jen Tsai6,7* 1 Institute of Biomedical Sciences, China Medical University, Taichung, Taiwan, 2 Vita Genomics, Inc., Taipei, Taiwan, 3 TickleFish Systems Corporation, Seattle, Western Australia, United States of America, a1111111111 4 Department of Public Health, Institute of Epidemiology and Preventive Medicine, National Taiwan a1111111111 University, Taipei, Taiwan, 5 Center for Neuropsychiatric Research, National Health Research Institutes, a1111111111 Miaoli County, Taiwan, 6 Department of Psychiatry, Taipei Veterans General Hospital, Taipei, Taiwan, 7 Division of Psychiatry, National Yang-Ming University, Taipei, Taiwan, 8 Department of Agronomy, College a1111111111 of Agriculture & Natural Resources, National Chung Hsing University, Taichung, Taiwan a1111111111 * [email protected] (EL); [email protected] (SJT) Abstract OPEN ACCESS Citation: Lin E, Kuo P-H, Liu Y-L, Yang AC, Kao C- Increased risk of developing metabolic syndrome (MetS) has been associated with the cir- F, Tsai S-J (2017) Effects of circadian clock genes cadian clock genes. In this study, we assessed whether 29 circadian clock-related genes and health-related behavior on metabolic (including ADCYAP1, ARNTL, ARNTL2, BHLHE40, CLOCK, CRY1, CRY2, CSNK1D, syndrome in a Taiwanese population: Evidence from association and interaction analysis. PLoS CSNK1E, GSK3B, HCRTR2, KLF10, NFIL3, NPAS2, NR1D1, NR1D2, PER1, PER2, ONE 12(3): e0173861. https://doi.org/10.1371/ PER3, REV1, RORA, RORB, RORC, SENP3, SERPINE1, TIMELESS, TIPIN, VIP, and journal.pone.0173861 VIPR2) are associated with MetS and its individual components independently and/or Editor: Etienne Challet, CNRS, University of through complex interactions in a Taiwanese population. -
Figure S1. Reverse Transcription‑Quantitative PCR Analysis of ETV5 Mrna Expression Levels in Parental and ETV5 Stable Transfectants
Figure S1. Reverse transcription‑quantitative PCR analysis of ETV5 mRNA expression levels in parental and ETV5 stable transfectants. (A) Hec1a and Hec1a‑ETV5 EC cell lines; (B) Ishikawa and Ishikawa‑ETV5 EC cell lines. **P<0.005, unpaired Student's t‑test. EC, endometrial cancer; ETV5, ETS variant transcription factor 5. Figure S2. Survival analysis of sample clusters 1‑4. Kaplan Meier graphs for (A) recurrence‑free and (B) overall survival. Survival curves were constructed using the Kaplan‑Meier method, and differences between sample cluster curves were analyzed by log‑rank test. Figure S3. ROC analysis of hub genes. For each gene, ROC curve (left) and mRNA expression levels (right) in control (n=35) and tumor (n=545) samples from The Cancer Genome Atlas Uterine Corpus Endometrioid Cancer cohort are shown. mRNA levels are expressed as Log2(x+1), where ‘x’ is the RSEM normalized expression value. ROC, receiver operating characteristic. Table SI. Clinicopathological characteristics of the GSE17025 dataset. Characteristic n % Atrophic endometrium 12 (postmenopausal) (Control group) Tumor stage I 91 100 Histology Endometrioid adenocarcinoma 79 86.81 Papillary serous 12 13.19 Histological grade Grade 1 30 32.97 Grade 2 36 39.56 Grade 3 25 27.47 Myometrial invasiona Superficial (<50%) 67 74.44 Deep (>50%) 23 25.56 aMyometrial invasion information was available for 90 of 91 tumor samples. Table SII. Clinicopathological characteristics of The Cancer Genome Atlas Uterine Corpus Endometrioid Cancer dataset. Characteristic n % Solid tissue normal 16 Tumor samples Stagea I 226 68.278 II 19 5.740 III 70 21.148 IV 16 4.834 Histology Endometrioid 271 81.381 Mixed 10 3.003 Serous 52 15.616 Histological grade Grade 1 78 23.423 Grade 2 91 27.327 Grade 3 164 49.249 Molecular subtypeb POLE 17 7.328 MSI 65 28.017 CN Low 90 38.793 CN High 60 25.862 CN, copy number; MSI, microsatellite instability; POLE, DNA polymerase ε. -

Grimme, Acadia.Pdf
MECHANISM OF ACTION OF HISTONE DEACETYLASE INHIBITORS ON SURVIVAL MOTOR NEURON 2 PROMOTER by Acadia L. Grimme A thesis submitted to the Faculty of the University of Delaware in partial fulfillment of the requirements for the degree of Bachelors of Science in Biological Sciences with Distinction Spring 2018 © 2018 Acadia Grimme All Rights Reserved MECHANISM OF ACTION OF HISTONE DEACETYLASE INHIBITORS ON SURVIVAL MOTOR NEURON 2 PROMOTER by Acadia L. Grimme Approved: __________________________________________________________ Matthew E. R. Butchbach, Ph.D. Professor in charge of thesis on behalf of the Advisory Committee Approved: __________________________________________________________ Deni S. Galileo, Ph.D. Professor in charge of thesis on behalf of the Advisory Committee Approved: __________________________________________________________ Carlton R. Cooper, Ph.D. Committee member from the Department of Biological Sciences Approved: __________________________________________________________ Gary H. Laverty, Ph.D. Committee member from the Board of Senior Thesis Readers Approved: __________________________________________________________ Michael Chajes, Ph.D. Chair of the University Committee on Student and Faculty Honors ACKNOWLEDGMENTS I would like to acknowledge my thesis director Dr. Butchbach for his wonderful guidance and patience as I worked through my project. He has been an excellent research mentor over the last two years and I am forever thankful that he welcomed me into his lab. His dedication to his work inspires me as an aspiring research scientist. His lessons will carry on with me as I pursue future research in graduate school and beyond. I would like to thank both current and former members of the Motor Neuron Disease Laboratory: Sambee Kanda, Kyle Hinkle, and Andrew Connell. Sambee and Andrew patiently taught me many of the techniques I utilized in my project, and without them it would not be what it is today. -
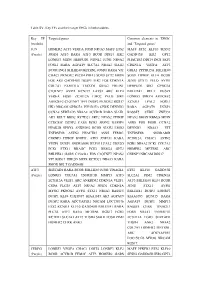
Targeted Genes Common Elements In
Table SV. Key TFs and their target DEGs in hub modules. Key TF Targeted genes Common elements in ‘DEGs’ (module) and ‘Targeted genes’ JUN HOMER2 ATF3 VEGFA FOSB NR4A3 MAFF ETS2 MAFF ETS2 KLF10 SESN2 (Purple) JOSD1 ATF3 RARA ATF3 BCOR DDIT4 IER2 GADD45B IER2 GPT2 LONRF3 MIDN HERPUD1 NDNL2 JUNB NR4A2 PHACTR3 DDIT4 ING1 SKP2 FOSL1 RARA AGPAT9 SLC7A1 NR4A2 SIAH2 CDKN1A VEGFA ATF3 BCOR ING1 BHLHE40 METRNL JOSD1 RARA VIT GRIA2 PPP1R15A BHLHE40 CHAC1 PKNOX2 PLCB4 PHF13 SOX9 SYT2 MIDN SOX9 ITPRIP KLF4 BCOR FOS AK5 GADD45B DUSP1 IER2 FOS CDKN1A JUNB STX11 PELO AVPI1 COL7A1 FAM131A TBCCD1 GRIA2 ERLIN1 HERPUD1 SIK1 GPRC5A C1QTNF7 AVPI1 KCNJ15 LATS2 ARC KLF4 BHLHE41 RELT DUSP2 VEGFA FOSB ZC3H12A LMO2 PELO SIK1 LONRF3 SPRY4 ARHGEF2 ARHGEF2 C1QTNF7 TFPI DUSP2 PKNOX2 RGS17 KCNJ15 LPAL2 FOSL1 HK1 NRCAM GPRC5A PPP1R15A CERK DENND3 RARA AGPAT9 DUSP1 CCNA2 SERTAD3 NR4A2 ACVR1B RARA SIAH1 RASSF5 CERK ZNF331 AK5 RELT BRD2 KCTD21 SKP2 NPAS2 ITPRIP NPAS2 SMOX RBM24 MIDN CCDC85C DUSP2 CARS EGR1 SESN2 RASSF5 ASNS FOS FOSB CCNA2 PIP4K2B SPRY4 ANKRD52 BCOR SIAH2 LMO2 DENND3 NR4A3 VIT TNFRSF1B ASTN2 PHACTR3 ASNS FEM1C TNFRSF1B SNORA80B CSRNP3 ITPRIP BNIP3L ATF3 ZNF331 RARA ZC3H12A CHAC1 ASTN2 VEZF1 DUSP1 SNORA80B KLF10 LPAL2 UBE2O EGR1 NR4A2 PCK1 COL7A1 PCK1 STX11 RRAGC PCK1 RBM24 GPT2 HOMER2 METRNL ARC BHLHE41 GARS C15orf41 FOS C1QTNF7 NPAS2 CSRNP3 NRCAM RGS17 VIT RGS17 UBE2O SOX9 KCTD21 NR4A3 RARA SMOX RELT GADD45B ATF3 SERTAD1 RARA BCOR BHLHE40 JUNB TINAGL1 ETS2 KLF10 GADD45B (Purple) LONRF3 COL7A1 TNFRSF1B MMP13 ATF3 SLC2A1 PIM2 CDKN1A ZC3H12A VEZF1 ARC ANKRD52 -

(A) Heat Maps Showing Significantly Elevated Levels of Tgfβ/SMAD Pathway-Related Genes In
Supplemental Figure and Legends Supplemental Figure 1. PLIN2KD significantly altered TGFβ/SMAD pathway genes. (A) Heat maps showing significantly elevated levels of TGFβ/SMAD pathway-related genes in PLIN2KD EndoCβH2-Cre β cells. The color keys represent the row-wise Z-score. (B) Notably, GDF15 level appears to be PLIN2 dependent (directly extracted from bulk RNA-seq data), TPM: Transcripts Per Million. All error bars indicate SD, n=3, * P<0.05 vs Sham. Supplemental Figure 2. NAC cannot rescue EA induced effectors of ER stress. EndoCβH2-Cre cells (termed Sham) were treated with EA (500 µM, 24h) in the presence of absence of N-acetyl cysteine (NAC, 1mM). The expression of stress induced IRE1 and XBP-1 ratio (i.e., spliced/total) mRNA levels were analyzed by qPCR. All error bars indicate SD, n=3, * P<0.05, ** P<0.01, *** P<0.05 vs Sham Supplemental Figure 3. EA provoked a stronger stress gene response than PA. Sham cells were treated with PA (500 µM, 48 hours) or EA (500 µM, 24 hours) and then analyzed by qRT-PCR for FEV, IRE1 and XBP-1 ratio (i.e., spliced/total). All of these PLIN2KD and ER stress markers were more highly elevated in EA cells. All error bars indicate SD, n=3, * P<0.05, ** P<0.01, *** P<0.05 vs Sham. Supplemental Figure 4. PLIN2 manipulation induced many changes in the expression of gene important to mitochondria function and health. Heat maps showing changes in many different mitochondrial genes in PLIN2KD and PLIN2OE cells, including (A) electron transport chain and mitochondrial encoding, (B) fusion and fission, (C) ion balance and (D) mitophagy. -

Circadian- and Sex-Dependent Increases in Intravenous Cocaine Self-Administration in Npas2 Mutant Mice
Research Articles: Behavioral/Cognitive Circadian- and sex-dependent increases in intravenous cocaine self-administration in Npas2 mutant mice https://doi.org/10.1523/JNEUROSCI.1830-20.2020 Cite as: J. Neurosci 2020; 10.1523/JNEUROSCI.1830-20.2020 Received: 15 July 2020 Revised: 13 November 2020 Accepted: 18 November 2020 This Early Release article has been peer-reviewed and accepted, but has not been through the composition and copyediting processes. The final version may differ slightly in style or formatting and will contain links to any extended data. Alerts: Sign up at www.jneurosci.org/alerts to receive customized email alerts when the fully formatted version of this article is published. Copyright © 2020 the authors 1 Circadian- and sex-dependent increases in intravenous cocaine self-administration in Npas2 mutant mice 2 3 Abbreviated title: Increased cocaine intake in female Npas2 mutants 4 5 Lauren M. DePoy1,2, Darius D. Becker-Krail1,2, Wei Zong3, Kaitlyn Petersen1,2, Neha M. Shah1, Jessica H. 6 Brandon1, Alyssa M. Miguelino1, George C. Tseng3, Ryan W. Logan1,2,4, *Colleen A. McClung1,2,4 7 8 1Department of Psychiatry, Translational Neuroscience Program, University of Pittsburgh School of Medicine, 9 15219 10 2Center for Neuroscience, University of Pittsburgh, 15261 11 3Department of Biostatistics, University of Pittsburgh, 15261 12 4Center for Systems Neurogenetics of Addiction, The Jackson Laboratory, 04609 13 14 15 *Corresponding Author: 16 Colleen A. McClung, PhD 17 450 Technology Dr. Ste 223 18 Pittsburgh, PA 15219 19 412-624-5547 20 [email protected] 21 22 Key words: circadian, sex-differences, cocaine, self-administration, substance use, Npas2 23 24 Pages: 37 25 Figures: 9 26 Extended data tables: 3 27 Abstract: 250 28 Introduction: 552 29 Discussion: 1490 30 31 Disclosures: 32 All authors have no financial disclosures or conflicts of interest to report. -

BHLHE41 Is Differentially Expressed in Multiple Models of Coronavirus Infection-PDF 032120
1 The transcription factor and basic helix-loop-helix family member e41 BHLHE41 is differentially expressed and transcriptionally induced in models of coronavirus infection. 2 1 3 Shahan Mamoor 1Thomas Jefferson School of Law 4 San Diego, CA 92101 [email protected] 5 6 The coronavirus COVID19 pandemic is an emerging biosafety threat to the nation and the world (1). There are no treatments approved for coronavirus infection in humans (2) and there 7 is a lack of information available regarding the basic transcriptional behavior of human cells 8 and mammalian tissues following coronavirus infection. We mined two independent datasets (3, 4), public (3) and published (4) containing transcriptome data from infection models of the 9 Middle East respiratory syndrome (MERS) coronavirus and human coronavirus (HCoV) to discover genes that are differentially expressed in coronaviruses and identify potential 10 therapeutic targets and host cell vulnerabilities. We identified the transcription factor basic helix-loop-helix family member e41 BHLHE41 as a conserved differentially expressed gene 11 following coronavirus infection. BHLHE41 may be involved in the cellular response to COVID19 infection. 12 13 14 15 16 17 18 19 20 21 22 23 24 25 Keywords: BHLHE41, coronavirus, MERS coronavirus, human coronavirus, HCoV, systems 26 biology of viral infection, COVID19. 27 28 1 1 Viruses are classified according to a system known as the “Baltimore” classification of 2 viruses (5) wherein the characteristics of the viral genome, including whether it is plus or minus 3 strand, also known as positive-sense or negative-sense, whether the genome is single- 4 stranded or double-stranded, whether their genome is composed or RNA or DNA, and whether 5 or not they use a reverse transcriptase to replicate are used to group viruses. -

Expression Profiling Associates Blood and Brain Glucocorticoid Receptor
Expression profiling associates blood and brain SEE COMMENTARY glucocorticoid receptor signaling with trauma-related individual differences in both sexes Nikolaos P. Daskalakisa,b,1, Hagit Cohenc, Guiqing Caia,d, Joseph D. Buxbauma,d,e, and Rachel Yehudaa,b,e Departments of aPsychiatry, dGenetics and Genomic Sciences, and eNeuroscience, Icahn School of Medicine at Mount Sinai, New York, NY 10029; bMental Health Patient Care Center, James J. Peters Veterans Affairs Medical Center, Bronx, NY 10468; and cAnxiety and Stress Research Unit, Ministry of Health Mental Health Center, Faculty of Health Sciences, Ben-Gurion University of the Negev, Beer Sheva 84170, Israel Edited by Bruce S. McEwen, The Rockefeller University, New York, NY, and approved July 14, 2014 (received for review February 7, 2014) Delineating the molecular basis of individual differences in the stress response to stress (4, 5). The emergence of system- and genome- response is critical to understanding the pathophysiology and treat- wide approaches permits the opportunity for unbiased identifi- ment of posttraumatic stress disorder (PTSD). In this study, 7 d after cation of novel pathways. Because PTSD is more prevalent in predator-scent-stress (PSS) exposure, male and female rats were women than men (1), and sex is a potential source of response classified into vulnerable (i.e., “PTSD-like”) and resilient (i.e., minimally variation to trauma in both animals (6) and humans (7), it is also affected) phenotypes on the basis of their performance on a variety of critical to include both sexes in such studies. behavioral measures. Genome-wide expression profiling in blood and In the present study, PSS-exposed male and female rats were two limbic brain regions (amygdala and hippocampus), followed by behaviorally tested in EPM and ASR tests a week after PSS and divided in EBR and MBR groups [at this point, the behavioral quantitative PCR validation, was performed in these two groups of response of the rats is stable in terms of prevalence of EBRs vs.