Fusing Catalase to an Alkane-Producing Enzyme
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Biomolecules
CHAPTER 3 Biomolecules 3.1 Carbohydrates In the previous chapter you have learnt about the cell and 3.2 Fatty Acids and its organelles. Each organelle has distinct structure and Lipids therefore performs different function. For example, cell membrane is made up of lipids and proteins. Cell wall is 3.3 Amino Acids made up of carbohydrates. Chromosomes are made up of 3.4 Protein Structure protein and nucleic acid, i.e., DNA and ribosomes are made 3.5 Nucleic Acids up of protein and nucleic acids, i.e., RNA. These ingredients of cellular organelles are also called macromolecules or biomolecules. There are four major types of biomolecules— carbohydrates, proteins, lipids and nucleic acids. Apart from being structural entities of the cell, these biomolecules play important functions in cellular processes. In this chapter you will study the structure and functions of these biomolecules. 3.1 CARBOHYDRATES Carbohydrates are one of the most abundant classes of biomolecules in nature and found widely distributed in all life forms. Chemically, they are aldehyde and ketone derivatives of the polyhydric alcohols. Major role of carbohydrates in living organisms is to function as a primary source of energy. These molecules also serve as energy stores, 2021-22 Chapter 3 Carbohydrade Final 30.018.2018.indd 50 11/14/2019 10:11:16 AM 51 BIOMOLECULES metabolic intermediates, and one of the major components of bacterial and plant cell wall. Also, these are part of DNA and RNA, which you will study later in this chapter. The cell walls of bacteria and plants are made up of polymers of carbohydrates. -

Aldehydes and Ketones
12 Aldehydes and Ketones Ethanol from alcoholic beverages is first metabolized to acetaldehyde before being broken down further in the body. The reactivity of the carbonyl group of acetaldehyde allows it to bind to proteins in the body, the products of which lead to tissue damage and organ disease. Inset: A model of acetaldehyde. (Novastock/ Stock Connection/Glow Images) KEY QUESTIONS 12.1 What Are Aldehydes and Ketones? 12.8 What Is Keto–Enol Tautomerism? 12.2 How Are Aldehydes and Ketones Named? 12.9 How Are Aldehydes and Ketones Oxidized? 12.3 What Are the Physical Properties of Aldehydes 12.10 How Are Aldehydes and Ketones Reduced? and Ketones? 12.4 What Is the Most Common Reaction Theme of HOW TO Aldehydes and Ketones? 12.1 How to Predict the Product of a Grignard Reaction 12.5 What Are Grignard Reagents, and How Do They 12.2 How to Determine the Reactants Used to React with Aldehydes and Ketones? Synthesize a Hemiacetal or Acetal 12.6 What Are Hemiacetals and Acetals? 12.7 How Do Aldehydes and Ketones React with CHEMICAL CONNECTIONS Ammonia and Amines? 12A A Green Synthesis of Adipic Acid IN THIS AND several of the following chapters, we study the physical and chemical properties of compounds containing the carbonyl group, C O. Because this group is the functional group of aldehydes, ketones, and carboxylic acids and their derivatives, it is one of the most important functional groups in organic chemistry and in the chemistry of biological systems. The chemical properties of the carbonyl group are straightforward, and an understanding of its characteristic reaction themes leads very quickly to an understanding of a wide variety of organic reactions. -

Aldehydes and Ketones Are Simple Organic Compounds Containing a Carbonyl Group
Aldehydes and Ketones are simple organic compounds containing a carbonyl group. Carbonyl group contains carbon- oxygen double bond. These organic compounds are simple because the carbon atom presents in the carbonyl group lack reactive groups such as OH or Cl. By Dr. Sayed Hasan Mehdi Assistant Professor Department of Chemistry Shia P.G. College, Lucknow Dr. S.Hasan Mehdi 6/13/2020 This is to bring to kind notice that the matter of this e- content is for the B.Sc. IV semester students. It has been taken from the following sources. The students are advised to follow these books as well. •A TEXTBOOK OF ORGANIC CHEMISTRY by Arun Bahl & B.S. Bahl, S. Chand & Company Ltd. Publication. •Graduate Organic Chemistry by M. K. Jain and S.C. Sharma, Vishal Publishing Co. •Pradeep’s Organic Chemistry Vol II by R. N. Dhawan, Pradeep Publication, Jalandhar. Dr. S.Hasan Mehdi 6/13/2020 An aldehyde is one of the classes of carbonyl group- containing alkyl group on one end and hydrogen on the other end. The R and Ar denote alkyl or aryl member respectively. In the condensed form, the aldehyde is written as –CHO. Dr. S.Hasan Mehdi 6/13/2020 Dr. S.Hasan Mehdi 6/13/2020 1. From Alcohols: a. By oxidation of Alcohols: Aldehydes and ketones can be prepared by the controlled oxidation of primary and secondary alcohols using an acidified solution of potassium dichromate or permanganate. Primary alcohol produces aldehydesRef. Last slide. O K Cr O RCH2OH + [O] 2 2 7 + R C H H 10 Alcohol Aldehyde O CH3CH2OH+ [O] K2Cr2O7 + CH3 C H H Ethyl Alcohol Acetaldehyde The aldehydes formed in the above reaction are very easily oxidised to carboxylc acids if allowed to remain in the reaction mixture. -

Sugars As the Source of Energized Carbon for Abiogenesis
Astrobiology Science Conference 2010 (2010) 5095.pdf SUGARS AS THE SOURCE OF ENERGIZED CARBON FOR ABIOGENESIS. A. L. Weber, SETI Institute, NASA Ames Research Center, Mail Stop 239-4, Moffett Field, CA, 94035-1000, [email protected] Abstract: As shown in Figure 1, abiogenesis has sev- eral requirements: (A) a source of organic substrates and chemical energy that drives the synthesis of (B) useful small molecules (ammonia, monomers, metabo- lites, energy molecules), and (C) a second synthetic processs that yields large replicating and catalytic polymers that control (D) the growth and maintenance of a primitive protocell. Furthermore, the required chemical energy must be sustained and effectively coupled to individual reactions to drive biosynthesis at a rate that counters chemical degradation. Energy coupling would have been especially difficult during the origin of life before the development of powerful enzyme catalysts with 3-D active sites. To solve this energy coupling problem we have investigated abio- genesis using sugar substrates whose energized carbon groups drive spontaneous synthetic self-transformation reactions that yield: biometabolites, catalytic mole- cules, energy-rich thioesters, amino acids, plausible alternative nucleobases and cell-like microstructures [1-8]. Recently, we demonstrated that sugars drive the synthesis of ammonia from nitrite [9]. The ability of sugars to drive ammonia synthesis provides a way to generate ammonia at microscopic sites of sugar-based origins processes, thereby eliminating the need for a planet-wide source of photochemically unstable am- monia. Figure 1. Major Synthetic Processes of Abiogenesis. [1] Weber A. L. (1998) Orig. Life Evol. Biosph., 28, 259-270. [2] Weber A. -

An Overview of Biosynthesis Pathways – Inspiration for Pharmaceutical and Agrochemical Discovery
An Overview of Biosynthesis Pathways – Inspiration for Pharmaceutical and Agrochemical Discovery Alan C. Spivey [email protected] 19th Oct 2019 Lessons in Synthesis - Azadirachtin • Azadirachtin is a potent insect anti-feedant from the Indian neem tree: – exact biogenesis unknown but certainly via steroid modification: O MeO C OAc O 2 H O OH O H O OH 12 O O C 11 O 14 OH oxidative 8 O H 7 cleavage highly hindered C-C bond HO OH AcO OH AcO OH for synthesis! H H of C ring H MeO2C O AcO H tirucallol azadirachtanin A azadirachtin (cf. lanosterol) (a limanoid = tetra-nor-triterpenoid) – Intense synhtetic efforts by the groups of Nicolaou, Watanabe, Ley and others since structural elucidation in 1987. –1st total synthesis achieved in 2007 by Ley following 22 yrs of effort – ~40 researchers and over 100 person-years of research! – 64-step synthesis – Veitch Angew. Chem. Int. Ed. 2007, 46, 7629 (DOI) & Veitch Angew. Chem. Int. Ed. 2007, 46, 7633 (DOI) – Review ‘The azadirachtin story’ see: Veitch Angew. Chem. Int. Ed. 2008, 47, 9402 (DOI) Format & Scope of Presentation • Metabolism & Biosynthesis – some definitions, 1° & 2° metabolites • Shikimate Metabolites – photosynthesis & glycolysis → shikimate formation → shikimate metabolites – Glyphosate – a non-selective herbicide • Alkaloids – acetylCoA & the citric acid cycle → -amino acids → alkaloids – Opioids – powerful pain killers • Fatty Acids and Polyketides –acetylCoA → malonylCoA → fatty acids, prostaglandins, polyketides, macrolide antibiotics – NSAIDs – anti-inflammatory’s • Isoprenoids/terpenes -

Chapter 14 – Aldehydes and Ketones
Chapter 14 – Aldehydes and Ketones 14.1 Structures and Physical Properties of Aldehydes and Ketones Ketones and aldehydes are related in that they each possess a C=O (carbonyl) group. They differ in that the carbonyl carbon in ketones is bound to two carbon atoms (RCOR’), while that in aldehydes is bound to at least one hydrogen (H2CO and RCHO). Thus aldehydes always place the carbonyl group on a terminal (end) carbon, while the carbonyl group in ketones is always internal. Some common examples include (common name in parentheses): O O H HH methanal (formaldehyde) trans-3-phenyl-2-propenal (cinnamaldehyde) preservative oil of cinnamon O O propanone (acetone) 3-methylcyclopentadecanone (muscone) nail polish remover a component of one type of musk oil Simple aldehydes (e.g. formaldehyde) typically have an unpleasant, irritating odor. Aldehydes adjacent to a string of double bonds (e.g. 3-phenyl-2-propenal) frequently have pleasant odors. Other examples include the primary flavoring agents in oil of bitter almond (Ph- CHO) and vanilla (C6H3(OH)(OCH3)(CHO)). As your book says, simple ketones have distinctive odors (similar to acetone) that are typically not unpleasant in low doses. Like aldehydes, placing a collection of double bonds adjacent to a ketone carbonyl generally makes the substance more fragrant. The primary flavoring agent in oil of caraway is just a such a ketone. 2 O oil of carraway Because the C=O group is polar, small aldehydes and ketones enjoy significant water solubility. They are also quite soluble in typical organic solvents. 14.2 Naming Aldehydes and Ketones Aldehydes The IUPAC names for aldehydes are obtained by using rules similar to those we’ve seen for other functional groups (e.g. -

Matrix-Assisted Laser Desorption/Ionization Time-Of-Flight
Received: 26 November 2018 Revised: 29 January 2019 Accepted: 31 January 2019 DOI: 10.1002/rcm.8406 RESEARCH ARTICLE Matrix‐assisted laser desorption/ionization time‐of‐flight mass spectrometry analysis for characterization of lignin oligomers using cationization techniques and 2,5‐dihydroxyacetophenone (DHAP) matrix Amber S. Bowman | Shardrack O. Asare | Bert C. Lynn Department of Chemistry, University of Rationale: Effective analytical techniques are needed to characterize lignin Kentucky, Lexington, KY 40506, USA products for the generation of renewable carbon sources. Application of matrix‐ Correspondence assisted laser desorption/ionization (MALDI) in lignin analysis is limited because of Bert C. Lynn, Department of Chemistry, UK Mass Spectrometry Facility, University of poor ionization efficiency. In this study, we explored the potential of cationization Kentucky, A053 ASTeCC Building, Lexington, along with a 2,5‐dihydroxyacetophenone (DHAP) matrix to characterize model KY 40506‐0286, USA. Email: [email protected] lignin oligomers. Funding information Methods: Synthesized lignin oligomers were analyzed using the developed MALDI National Science Foundation, Grant/Award method. Two matrix systems, DHAP and α‐cyano‐4‐hydroxycinnamic acid (CHCA), Number: OIA 1632854 and three cations (lithium, sodium, silver) were evaluated using a Bruker UltraFlextreme time‐of‐flight mass spectrometer. Instrumental parameters, cation concentration, matrix, sample concentrations, and sample spotting protocols were optimized for improved results. Results: The DHAP/Li+ combination was effective for dimer analysis as lithium adducts. Spectra from DHP and ferric chloride oligomers showed improved signal intensities up to decamers (m/z 1823 for the FeCl3 system) and provided insights into differences in the oligomerization mechanism. Spectra from a mixed DHP oligomer system containing H, G, and S units showed contributions from all monolignols within an oligomer level (e.g. -

Particulate Preparation from Pisum Sativum (Plant Wax/CO Production/Hydrocarbon) T
Proc. Natl. Acad. Sci. USA Vol. 81, pp. 6613-6617, November 1984 Biochemistry Alkane biosynthesis by decarbonylation of aldehydes catalyzed by a particulate preparation from Pisum sativum (plant wax/CO production/hydrocarbon) T. M. CHEESBROUGH AND P. E. KOLATTUKUDY* Institute of Biological Chemistry and Biochemistry/Biophysics Program, Washington State University. Pullman, WA 99164-6340 Communicated by P. K. Stumpf, June 29, 1984 ABSTRACT Mechanism of enzymatic conversion of a fat- MATERIALS AND METHODS ty acid to the corresponding alkane by the loss of the carboxyl Materials. Omnifluor, [1-_4C]stearic acid, [U-3H]tetraco- carbon was investigated with particulate preparations from Pi- sanoic acid, [1-'4Cjsteroyl-CoA, and LiAl3H4 were from sum sativum. A heavy particulate preparation (sp. gr., 1.30 New England Nuclear. Sodium [1-14C]cyanide was from g/cm3) isolated by two density-gradient centrifugation steps ICN (Chemical and Radioisotopes Division). Pyridinium catalyzed conversion of octadecanal to heptadecane and CO. chlorochromate, meso-tetraphenylporphyrin, and RhCP Experiments with [1-3H,1_14C]octadecanal showed the stoichi- 3H2O were from Aldrich. LiAIH4, di-t-butylchlorophos- ometry of the reaction and retention of the aldehydic hydrogen phine, and Ru(CO)12 were from Alfa Products. The rhodium in the alkane during this enzymatic decarbonylation. This de- chelate was synthesized by the procedure of Monson (8). carbonylase showed an optimal pH of 7.0 and a Km of 35 ,uM Ruthenium dicarbonyl tetraphenylporphyrin [Ru(CO)2- for the aldehyde. This enzyme was severely inhibited by metal (TTP)] was synthesized (9) and activated by substitution of ion chelators and showed no requirement for any cofactors. -

Aldehydes Can React with Alcohols to Form Hemiacetals
340 14 . Nucleophilic substitution at C=O with loss of carbonyl oxygen You have, in fact, already met some reactions in which the carbonyl oxygen atom can be lost, but you probably didn’t notice at the time. The equilibrium between an aldehyde or ketone and its hydrate (p. 000) is one such reaction. O HO OH H2O + R1 R2 R1 R2 When the hydrate reverts to starting materials, either of its two oxygen atoms must leave: one OPh came from the water and one from the carbonyl group, so 50% of the time the oxygen atom that belonged to the carbonyl group will be lost. Usually, this is of no consequence, but it can be useful. O For example, in 1968 some chemists studying the reactions that take place inside mass spectrometers needed to label the carbonyl oxygen atom of this ketone with the isotope 18 O. 16 18 By stirring the ‘normal’ O compound with a large excess of isotopically labelled water, H 2 O, for a few hours in the presence of a drop of acid they were able to make the required labelled com- í In Chapter 13 we saw this way of pound. Without the acid catalyst, the exchange is very slow. Acid catalysis speeds the reaction up by making a reaction go faster by raising making the carbonyl group more electrophilic so that equilibrium is reached more quickly. The the energy of the starting material. We 18 also saw that the position of an equilibrium is controlled by mass action— O is in large excess. -

Synthesis and Biosynthesis of Polyketide Natural Products
Syracuse University SURFACE Chemistry - Dissertations College of Arts and Sciences 12-2011 Synthesis and Biosynthesis of Polyketide Natural Products Atahualpa Pinto Syracuse University Follow this and additional works at: https://surface.syr.edu/che_etd Part of the Chemistry Commons Recommended Citation Pinto, Atahualpa, "Synthesis and Biosynthesis of Polyketide Natural Products" (2011). Chemistry - Dissertations. 181. https://surface.syr.edu/che_etd/181 This Dissertation is brought to you for free and open access by the College of Arts and Sciences at SURFACE. It has been accepted for inclusion in Chemistry - Dissertations by an authorized administrator of SURFACE. For more information, please contact [email protected]. Abstract Traditionally separate disciplines of a large and broad chemical spectrum, synthetic organic chemistry and biochemistry have found in the last two decades a fertile common ground in the area pertaining to the biosynthesis of natural products. Both disciplines remain indispensable in providing unique solutions on numerous questions populating the field. Our contributions to this interdisciplinary pursuit have been confined to the biosynthesis of polyketides, a therapeutically and structurally diverse class of natural products, where we employed both synthetic chemistry and biochemical techniques to validate complex metabolic processes. One such example pertained to the uncertainty surrounding the regiochemistry of dehydration and cyclization in the biosynthetic pathway of the marine polyketide spiculoic acid A. The molecule's key intramolecular cyclization was proposed to occur through a linear chain containing an abnormally dehydrated polyene system. We synthesized a putative advanced polyketide intermediate and tested its viability to undergo a mild chemical transformation to spiculoic acid A. In addition, we applied a synthetic and biochemical approach to elucidate the biosynthetic details of thioesterase-catalyzed macrocyclizations in polyketide natural products. -
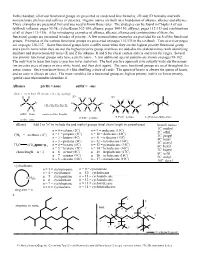
In This Handout, All of Our Functional Groups Are Presented As Condensed Line Formulas, 2D and 3D Formulas and with Nomenclature Prefixes and Suffixes (If Present)
In this handout, all of our functional groups are presented as condensed line formulas, 2D and 3D formulas and with nomenclature prefixes and suffixes (if present). Organic names are built on a foundation of alkanes, alkenes and alkynes. Those examples are presented first and you need to know those rules. The strategies can be found in Chapter 4 of our textbook (alkanes: pages 93-98, cycloalkanes 102-104, alkenes: pages 104-110, alkynes: pages 112-113 and combinations of all of them 113-115). After introducing examples of alkanes, alkenes, alkynes and combinations of them, the functional groups are presented in order of priority. A few nomenclature examples are provided for each of the functional groups. Examples of the various functional groups are presented on pages 115-135 in the textbook. Two overview pages are on pages 136-137. Some functional groups have a suffix name when they are the highest priority functional group and a prefix name when they are not the highest priority group, and these are added to the skeletal names with identifying numbers and stereochemistry terms (E and Z for alkenes, R and S for chiral centers and cis and trans for rings). Several low priority functional groups only have a prefix name. A few additional special patterns are shown on pages 98-102. The only way to learn this topic is practice (over and over). The best practice approach is to actually write out the names (on an extra piece of paper or on a white board, and then do it again). The same functional groups are used throughout the entire course. -

19.1 Ketones and Aldehydes
19.1 Ketones and Aldehydes • Both functional groups possess the carbonyl group • Important in both biology and industry Simplest aldehyde Simplest ketone used as a used mainly as preservative a solvent Copyright © 2017 John Wiley & Sons, Inc. All rights reserved. 19-1 Klein, Organic Chemistry 3e 19.1 Ketones And Aldehydes Copyright © 2017 John Wiley & Sons, Inc. All rights reserved. 19-2 Klein, Organic Chemistry 3e 19.2 Nomenclature • Four discrete steps to naming an aldehyde or ketone • Same procedure as with alkanes, alcohols, etc… 1. Identify and name the parent chain 2. Identify the name of the substituents (side groups) 3. Assign a locant (number) to each substituents 4. Assemble the name alphabetically Copyright © 2017 John Wiley & Sons, Inc. All rights reserved. 19-3 Klein, Organic Chemistry 3e 19.2 Nomenclature 1. Identify and name the parent chain – For aldehydes, replace the “-e” ending with an “-al” – the parent chain must include the carbonyl carbon Copyright © 2017 John Wiley & Sons, Inc. All rights reserved. 19-4 Klein, Organic Chemistry 3e 19.2 Nomenclature 1. Identify and name the parent chain – The aldehydic carbon is assigned number 1: Copyright © 2017 John Wiley & Sons, Inc. All rights reserved. 19-5 Klein, Organic Chemistry 3e 19.2 Nomenclature 1. Identify and name the parent chain – For ketones, replace the “-e” ending with an “-one” – The parent chain must include the C=O group – the C=O carbon is given the lowest #, and can be expressed before the parent name or before the suffix Copyright © 2017 John Wiley & Sons, Inc. All rights reserved.