Supplement to the Pre-Season Run
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Chilliwacklake Mp
M ANAGEMENT LAN P July 2000 for Chilliwack Lake Park & Chilliwack River Ecological Ministry of Environment, Reserve Lands and Parks BC Parks Division Chilliwack Lake Provincial Park & Chilliwack River Ecological Reserve M ANAGEMENT LAN P Prepared by BC Parks Lower Mainland District North Vancouver BC Canadian Cataloguing in Publication Data BC Parks. Lower Mainland District. Chilliwack Lake Provincial Park & Chilliwack River Ecological Reserve management plan Cover title: Management plan for Chilliwack Lake Park & Chilliwack River Ecological Reserve. ISBN 0-7726-4357-1 1. Provincial parks and reserves - British Columbia - Planning. 2. Provincial parks and reserves - British Columbia - Management. 3. Ecological reserves - British Columbia - Planning. 4. Ecological reserves - British Columbia - Management. 5. Chilliwack Lake Park (B.C.) 6. Chilliwack River Ecological Reserve (B.C.) I. Title. II. Title: Management plan for Chilliwack Lake Park & Chilliwack River Ecological Reserve. FC3815.C45B32 2000 333.78'30971137 C00-960299-2 F1089.C45B32 2000 TABLE OF CONTENTS Introduction......................................................................................................................................1 The Management Planning Process.....................................................................................1 Relationship to Other Land Use Planning Initiatives ..........................................................1 Background Summary .........................................................................................................1 -
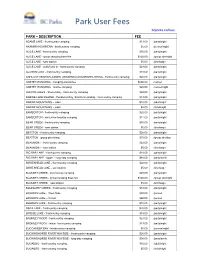
Park User Fees Bcparks.Ca/Fees
Park User Fees bcparks.ca/fees PARK – DESCRIPTION FEE ADAMS LAKE - frontcountry camping $13.00 /party/night AKAMINA-KISHINENA - backcountry camping $5.00 /person/night ALICE LAKE - frontcountry camping $35.00 /party/night ALICE LAKE - group camping base fee $120.00 /group site/night ALICE LAKE - sani station $5.00 /discharge ALICE LAKE - walk/cycle in - frontcountry camping $23.00 /party/night ALLISON LAKE - frontcountry camping $18.00 /party/night ANHLUUT’UKWSIM LAXMIHL ANGWINGA’ASANSKWHL NISGA - frontcountry camping $20.00 /party/night ANSTEY-HUNAKWA - camping-annual fee $600.00 /vessel ANSTEY-HUNAKWA - marine camping $20.00 /vessel/night ARROW LAKES - Shelter Bay - frontcountry camping $20.00 /party/night BABINE LAKE MARINE - Pendleton Bay, Smithers Landing - frontcountry camping $13.00 /party/night BABINE MOUNTAINS – cabin $10.00 /adult/night BABINE MOUNTAINS – cabin $5.00 /child/night BAMBERTON - frontcountry camping $20.00 /party/night BAMBERTON - winter frontcountry camping $11.00 /party/night BEAR CREEK - frontcountry camping $35.00 /party/night BEAR CREEK - sani station $5.00 /discharge BEATTON - frontcountry camping $20.00 /party/night BEATTON - group picnicking $35.00 /group site/day BEAUMONT - frontcountry camping $22.00 /party/night BEAUMONT - sani station $5.00 /discharge BIG BAR LAKE - frontcountry camping $18.00 /party/night BIG BAR LAKE - Upper - long-stay camping $88.00 /party/week BIRKENHEAD LAKE - frontcountry camping $22.00 /party/night BIRKENHEAD LAKE - sani station $5.00 /discharge BLANKET CREEK - frontcountry camping -

Download Download
Ames, Kenneth M. and Herbert D.G. Maschner 1999 Peoples of BIBLIOGRAPHY the Northwest Coast: Their Archaeology and Prehistory. Thames and Hudson, London. Abbas, Rizwaan 2014 Monitoring of Bell-hole Tests at Amoss, Pamela T. 1993 Hair of the Dog: Unravelling Pre-contact Archaeological Site DhRs-1 (Marpole Midden), Vancouver, BC. Coast Salish Social Stratification. In American Indian Linguistics Report on file, British Columbia Archaeology Branch, Victoria. and Ethnography in Honor of Lawrence C. Thompson, edited by Acheson, Steven 2009 Marpole Archaeological Site (DhRs-1) Anthony Mattina and Timothy Montler, pp. 3-35. University of Management Plan—A Proposal. Report on file, British Columbia Montana Occasional Papers No. 10, Missoula. Archaeology Branch, Victoria. Andrefsky, William, Jr. 2005 Lithics: Macroscopic Approaches to Acheson, S. and S. Riley 1976 Gulf of Georgia Archaeological Analysis (2nd edition). Cambridge University Press, New York. Survey: Powell River and Sechelt Regional Districts. Report on Angelbeck, Bill 2015 Survey and Excavation of Kwoiek Creek, file, British Columbia Archaeology Branch, Victoria. British Columbia. Report in preparation by Arrowstone Acheson, S. and S. Riley 1977 An Archaeological Resource Archaeology for Kanaka Bar Indian Band, and Innergex Inventory of the Northeast Gulf of Georgia Region. Report on file, Renewable Energy, Longueuil, Québec. British Columbia Archaeology Branch, Victoria. Angelbeck, Bill and Colin Grier 2012 Anarchism and the Adachi, Ken 1976 The Enemy That Never Was. McClelland & Archaeology of Anarchic Societies: Resistance to Centralization in Stewart, Toronto, Ontario. the Coast Salish Region of the Pacific Northwest Coast. Current Anthropology 53(5):547-587. Adams, Amanda 2003 Visions Cast on Stone: A Stylistic Analysis of the Petroglyphs of Gabriola Island, B.C. -

INITIAL ASSESSMENT of the CHILL PROPERTY New Westminster Mining Division Chilliwack Lake, BC for MAMMOTH GEOLOGICAL LTD. Bv
WMOTH COLOGICAL LTD. Box 5250, Port Hardy, B.C. VON 2PO Phone : (604) 949-5197 Fax : (604) 949-5197 INITIAL ASSESSMENT OF THE CHILL PROPERTY New Westminster Mining Division Chilliwack Lake, B.C. FOR MAMMOTH GEOLOGICAL LTD. Bv: R.Tim Henneberry, P.Geo. J December 7, l9<199 "I i -2- . SUMMARY The Chill property lies within the New Westminster Mining Division, east of Chilliwack at the north end of Chilliwack Lake. The property is being explored as a potential source of gran- ite dimension stone, for structural stone. The property was acquired as part of an on-going exploration and assessment program of the southern interior plutons. This claim covers the former Chilliwack Lake Quarry of Quarry Pacific Industries. Polished sections show the stone has an appealing texture and exhibits few flaws on the finished surface. Examination of existing quarry faces showed numerous flaws making the stone unsuitable for polished stone. The stone is suitable for split (structural) stone, exhibiting characteristics very similar to the light grey Hope Stone. Based on this observation, a & 500 ton bulk sampling program is recommended. The aim of this program will be to quarry and split 50 tons of stone and market the stone to have it specified on a number ofjob sites in the lower Mainland. The cost of this program is estimated at $43,183. The initial prospecting and mapping program completed on the Chill property cost $1,375. -3- . TABLE OF CONTENTS INTRODUCTION .............................................................................. .................................... -

Fieldnotes 2021-2022
FIELDNOTES 2021 – 2022 Pacific Science Field Operations Cover illustration: Copper Rockfish (Sebastes caurinus) in an old growth kelp forest covered in Proliferating Anemones (Epiactis prolifera). Queen Charlotte Strait, BC. Photo credit: Pauline Ridings, Fisheries and Oceans Canada. FIELDNOTES 2021 - 2022: DFO Pacific Science Field Operations TABLE OF CONTENT . INTRODUCTION 1 . DFO PACIFIC SCIENCE 2 . SCHEDULED FIELD OPERATIONS: 2021—2022 3 . DID YOU KNOW? 5 . REPORTING RESULTS 6 . ANNEX A PACIFIC SCIENCE ORGANIZATION 7 . ANNEX B FACT SHEET SERIES: 2020—2021 DFO Pacific Science Field Operations 12 . ANNEX C DATASETS PUBLISHED: 2020—2021 18 FIELDNOTES 2021 - 2022: DFO Pacific Science Field Operations INTRODUCTION Fisheries and Oceans Canada (DFO) conducts research and undertakes monitoring surveys of the marine and freshwater environment in support of sustainable fisheries, healthy aquatic ecosystems and living resources, and safe and effective marine services. In an effort to effectively inform and ensure Canadians feel engaged in the delivery of its science mandate, DFO produces Fieldnotes, an annual compendium of planned science field operations in the North Pacific and Arctic oceans, as well as in the coastal and interior waters of British Columbia and Yukon. Fieldnotes aims to: . inform Canadians of research and monitoring programming scheduled for the COVID-19 coming year; . promote the sharing of key information and data in a coordinated, timely, open and One year into the global pandemic, DFO transparent manner in order to encourage remains committed to delivering innovative dialogue and collaboration; science and services to Canadians. provide a platform from which to build and Following the suspension of scientific field nurture fundamentally more inclusive, trust- and respect-based relationships with all operations in the spring of 2020, DFO has Canadians; since resumed much of its field programming. -

Factors Limiting Juvenile Sockeye Production and Enhancement Potential for Selected B.C
Fisheries and Oceans Pêches et Océans Science Sciences C S A S S C C S Canadian Science Advisory Secretariat Secrétariat canadien de consultation scientifique Research Document 2001/098 Document de recherche 2001/098 Not to be cited without Ne pas citer sans permission of the authors 1 autorisation des auteurs 1 FACTORS LIMITING JUVENILE SOCKEYE PRODUCTION AND ENHANCEMENT POTENTIAL FOR SELECTED B.C. NURSERY LAKES K.S. Shortreed, K.F. Morton, K. Malange, and J.M.B. Hume Fisheries and Oceans Canada Marine Environment and Habitat Science Division 4222 Columbia Valley Highway Cultus Lake Laboratory, Cultus Lake, B.C. V2R 5B6 1 This series documents the scientific basis for 1 La présente série documente les bases the evaluation of fisheries resources in scientifiques des évaluations des ressources Canada. As such, it addresses the issues of halieutiques du Canada. Elle traite des the day in the time frames required and the problèmes courants selon les échéanciers documents it contains are not intended as dictés. Les documents qu’elle contient ne definitive statements on the subjects doivent pas être considérés comme des addressed but rather as progress reports on énoncés définitifs sur les sujets traités, mais ongoing investigations. plutôt comme des rapports d’étape sur les études en cours. Research documents are produced in the Les documents de recherche sont publiés dans official language in which they are provided to la langue officielle utilisée dans le manuscrit the Secretariat. envoyé au Secrétariat. This document is available on the Internet at: Ce document est disponible sur l’Internet à: http://www.dfo-mpo.gc.ca/csas/ ISSN 1480-4883 Ottawa, 2001 ABSTRACT In this report we present summaries of our current knowledge of freshwater factors limiting sockeye production from 60 B.C. -

British Columbia
BRITISH COLUMBIA RV PARKS & CAMPGROUNDS RECOMMENDED BY THE NRVOA British Columbia Canada's most westerly province, British Columbia is huge and diverse. If you love the great outdoors, you'll never be bored in a province that is so beautiful, with such variation, and with so many opportunities for hiking, rafting, sailing and skiing, to name just a few. British Columbia is studded with breathtakingly beautiful mountains that puncture vast, blue skies. You'll also find long stretches of rugged coastline, sandy beaches, wineries, orchards, forests and snowmelt-fed lakes. There is even Canada's only desert: Osoyoos, in the southern Okanagan, which is home to rattlesnakes, scorpions and prickly pear cacti. If you're seeking unspoiled wilderness, then British Columbia is the perfect province to lose yourself in. If you want company, however, there are plenty of modern and lively cities and towns in British Columbia. Vancouver has scores of galleries, museums and bars. But even in Vancouver you're never too removed from nature: the city contains Canada's largest city park, Stanley Park. Notable Attractions • Yoho National Park • Kootenay National Park • Tatshenshin-Alsek Provincial Park • Gwaii Haanas National Park Reserve & Haida Heritage Site • Banff National Park • Jasper National Park ...And Much More! Province Description Courtesy of iexplore Canada | NRVOA Recommended RV Parks & Campgrounds: 2019 Return to Table of Contents 2 TABLE OF CONTENTS 70 Mile House Sunset View Campground Alexis Creek Bull Canyon Campground Barkerville Bowron -

Scale Site SS Region SS District Site Name SS Location Phone
Scale SS SS Site Region District Site Name SS Location Phone 001 RCB DQU MISC SITES SIFR 01B RWC DQC ABFAM TEMP SITE SAME AS 1BB 2505574201 1001 ROM DPG BKB CEDAR Road past 4G3 on the old Lamming Ce 2505690096 1002 ROM DPG JOHN DUNCAN RESIDENCE 7750 Lower Mud river Road. 1003 RWC DCR PROBYN LOG LTD. Located at WFP Menzies#1 Scale Site 1004 RWC DCR MATCHLEE LTD PARTNERSHIP Tsowwin River estuary Tahsis Inlet 2502872120 1005 RSK DND TOMPKINS POST AND RAIL Across the street from old corwood 1006 RWC DNI CANADIAN OVERSEAS FOG CREEK - North side of King Isla 6046820425 1007 RKB DSE DYNAMIC WOOD PRODUCTS 1839 Brilliant Road Castlegar BC 2503653669 1008 RWC DCR ROBERT (ANDY) ANDERSEN Mobile Scale Site for use in marine 1009 ROM DPG DUNKLEY- LEASE OF SITE 411 BEAR LAKE Winton Bear lake site- Current Leas 2509984421 101 RWC DNI WESTERN FOREST PRODUCTS INC. MAHATTA RIVER (Quatsino Sound) - Lo 2502863767 1010 RWC DCR WESTERN FOREST PRODUCTS INC. STAFFORD Stafford Lake , end of Loughborough 2502863767 1011 RWC DSI LADYSMITH WFP VIRTUAL WEIGH SCALE Latitude 48 59' 57.79"N 2507204200 1012 RWC DNI BELLA COOLA RESOURCE SOCIETY (Bella Coola Community Forest) VIRT 2509822515 1013 RWC DSI L AND Y CUTTING EDGE MILL The old Duncan Valley Timber site o 2507151678 1014 RWC DNI INTERNATIONAL FOREST PRODUCTS LTD Sandal Bay - Water Scale. 2 out of 2502861881 1015 RWC DCR BRUCE EDWARD REYNOLDS Mobile Scale Site for use in marine 1016 RWC DSI MUD BAY COASTLAND VIRTUAL W/S Ladysmith virtual site 2507541962 1017 RWC DSI MUD BAY COASTLAND VIRTUAL W/S Coastland Virtual Weigh Scale at Mu 2507541962 1018 RTO DOS NORTH ENDERBY TIMBER Malakwa Scales 2508389668 1019 RWC DSI HAULBACK MILLYARD GALIANO 200 Haulback Road, DL 14 Galiano Is 102 RWC DNI PORT MCNEILL PORT MCNEILL 2502863767 1020 RWC DSI KURUCZ ROVING Roving, Port Alberni area 1021 RWC DNI INTERNATIONAL FOREST PRODUCTS LTD-DEAN 1 Dean Channel Heli Water Scale. -

Wilderness Trip Planner
National Park Service North Cascades National Park Service Complex U.S. Department of the Interior Stephen Mather Wilderness An Enduring Legacy of Wilderness “[I]t is hereby declared to be the policy of the Congress to secure for the American people of present and future generations the benefits of an enduring resource of wilderness.” — Wilderness Act, 1964 The North Cascades National Park Complex includes 684,000 acres in three units: North Cascades National Park, Lake Chelan National Recre- ation Area, and Ross Lake National Recreation Area. Congress has designated 94% of the Complex as the Stephen Mather Wilderness. Today, as in the past, wilderness is an important part of every American’s story. People seek out wilderness for a variety of reasons: physical or mental challenge; solitude, renewal, or a respite from modern life; or as a place to find inspiration and to explore our heritage. What draws you to visit wilderness? The Stephen Mather Wilderness is at the heart of over two million acres of some of the wildest lands remaining, a place “where the earth and its community of life are untrammeled by man….” Untrammeled (meaning“free of restraint,” “unconfined”) captures the essence of wilderness: a place where the natural processes of the land prevail, and the developments of modern technological society are substantially unnoticeable. Here, we are visitors, but we also come home—to our natural heritage. It is a place to experience our past, and a place to find future respite. This is the enduring legacy of wilderness. To Hope, B.C. -

Campings British Columbia
Campings British Columbia 100 Mile House en omgeving Bridal Falls/Rosedale - 100 Mile Motel & RV Park - Camperland RV Park - 100 Mile House Municipal Campground - Fraser Valley /Rainbow Ranch RV Park - Camp Bridal Anahim Lake - Escott Bay Resort Bridge Lake - Anahim Lake Resort & RV Park - Eagle Island Resort - Moosehaven Resort Argenta - Cottonwood Bay Resort - Kootenay Lake Provincial Park Burns Lake en omgeving Arras - Beaver Point Resort - Monkman Provincial Park - Burns Lake Village Campground - Ethel F. Wilson Memorial PP Barkerville - Babine Lake Marine PP - Pinkut Creek Site - Lowhee Campground - Babine Lake Marine PP– Pendleton Bay Site Barriere Cache Creek en omgeving - DeeJay RV Park and Campground - Historic Hat Creek Ranch - Brookside Campsite Bear Lake en omgeving - Ashcroft Legacy Park Campground in Ashcroft - Tudyah Lake Provincial Park - Crooked River Provincial Park Canal Flats - Whiskers Point Provincial Park - Whiteswan Lake Provincial Park Campground Big Lake Ranch Canim Lake - Horsefly Lake Provincial Park - Canim Lake Resort - Rainbow Resort Blue River - South Point Resort - Blue River Campground - Reynolds Resort Boston Bar Castlegar - Canyon Alpine RV Park & Campground - Castlegar RV Park & Campground - Blue Lake Resort - Kootenay River RV Park - Tuckkwiowhum Campground Chase Boswell - Niskonlith Lake Provincial Park - Lockhart Beach Provincial Park - Bayshore Resort Chilliwack en omgeving - Cottonwood RV Park - Vedder River Campground - Sunnyside Campground in Cultus Lake - Cultus Lake Provincial Park Christina -

Outdoor Recreation & Tourism in the Fraser Valley Regional District
Regional Snapshot Series: Parks and Recreation Outdoor Recreation & Tourism in the Fraser Valley Regional District TABLE OF CONTENTS Utilizing our Natural Assets Existing Opportunities A Growing Demand for Parks and Recreation Investing in Our Region Harrison Hot Springs Nature-Based Tourism Tourism Growth Sectors Experience the Fraser Keeping up with Demand Mount Cheam The Fraser Valley Regional District is comprised of six member municipalities and seven electoral areas. City of Abbotsford, City of Chilliwack, District of Mission, District of Hope, District of Kent, Village of Harrison Hot Springs and Electoral Areas A, B, C, D, E, F and G. Fraser Valley Regional District OUR NATURAL ASSETS OUTDOOR Recreation IN FVRD Incredible natural environment and recreation potential The Fraser Valley Regional District (FVRD) is comprised of six member municipalities and seven electoral areas and features a variety of diverse communities, from small rural hamlets to the fifth largest city in British Columbia. The equally diverse landscape of rugged mountains, sizeable rivers, and fertile valleys offers a myriad of recreational opportunities including hiking, swimming, camping, biking, rafting and fishing, all of Cheam Ridge Regional Trail which are made use of by residents of the FVRD and beyond. A growing population combined with an escalating interest in outdoor recreation is increasing the demand for outdoor recreational opportunities in the area: a trend that is likely to continue. This region’s incredible natural environment and recreation potential benefits the economy by attracting tourists, businesses and residents. Parks and open space provide wildlife habitat, ensure biodiversity, and mitigate the effects of climate change. In addition, parks are some of the best places to educate people about their natural surroundings and the impact they can have. -

Preliminary 2018 Salmon Outlook December 2017 Pg 1 of 28
Preliminary 2018 Salmon Outlook December 2017 pg 1 of 28 PRELIMINARY 2018 SALMON OUTLOOK Since 2002, Pacific Region (BC & Yukon) Stock Assessment staff has provided a categorical outlook for the next year’s salmon returns. The Outlook is intended to provide an objective and consistent context within which to initiate fisheries planning. In particular, it provides a preliminary indication of salmon production and associated fishing opportunities by geographic area and species stock groups called an Outlook Unit. The Conservation Units covered by each Outlook Unit are listed in Appendix 1. Final stock-specific fishing plans described in the annual Salmon Integrated Fisheries Management Plans (IFMP) may be different from the generic scenarios described here. Stock-specific plans are informed by available science and management information, the specific nature of fisheries on a given stock, allocation policy, consultation input and other considerations. Actual fishing opportunities are subject to in-season information and are announced in-season via fishery notice or other official communications from DFO. For each Outlook Unit, an Outlook Category is provided on a scale of 1 to 4 (table below). The category reflects the current interpretation of available quantitative and qualitative information, including pre-season forecasts if available, and the opinion of DFO Stock Assessment staff. Where management targets for stocks have not been formally described, interim targets were either based on historical return levels or, if necessary, opinion of local staff. In some cases, multiple categories are reported to reflect variation in status among component populations within the Outlook Unit, or to capture the degree of uncertainty in the assessment.