SH 10.? :A7 R65 I9b9 This Document Is Made Available Electronically by the Minnesota Legislative Reference Library As Part of an Ongoing Digital Archiving Project
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

What the “Trail Eyes” Pros Taught Us About the SHT P H
A publication oF the Superior Hiking TrAil AssoCiation SUmmEr 2019 What the “Trail Eyes” Pros Taught Us About the SHT P H o im Malzhan iS the trail operations director T o for our sister trail organization the ice Age B y Fr Trail Alliance in Wisconsin. Doing business as esh T “Trail Eyes,” Tim was one of four entities the SHTA Tr hired in the fall of 2018 to evaluate and recom- ac mend renewal strategies for what we have dubbed k S mE D “The Big Bad Five,” those sections of the SHT most damaged from heavy use and old age (or both). i A Though all four evaluators—malzhan, Critical Connections Ecological Services (Jason and Amy Husveth), the north Country Trail Association, and (Continued on page 2) What the “Trail Eyes” Taught Us About the SHT (continued from cover) Great Lakes Trail Builders (Wil- lie Bittner)—did what we asked (provide specific prescriptions for the Big Bad Five), their ex- pert observations gave us much more: they shed light on the en- tire Superior Hiking Trail. In other words, what they saw on the Split Rock River loop, or the sections from Britton Peak to Oberg Mountain and Oberg to the Lutsen ski complex, or the proposed reroute of the SHT north of Gooseberry Falls State Park, were microcosms of bigger, more systemic issues with the SHT. ❚ “keep people on the Trail and water off of it.” This suc- cinct wisdom comes from Matt no bridge is not the only problem at the Split rock river loop. -

Map 2, Lake Superior State Water Trail from Knife River to Split Rock
ROUTE DESCRIPTION - River miles 26 to 60 (34 miles) (0.0 at Minnesota Entrance – Duluth Lift Bridge). 48.0 Private resort. [47° 07.135' N / 91° 30.265' W] 57.7 Little Two Harbors at Split Rock Lighthouse State Park. Access to park and lighthouse, a MAP 2 - Knife River to Split Rock Lighthouse State Park 51.0 Gooseberry Falls State Park and Gooseberry Minnesota Historic Site. Trailer access, parking, River. Carry-in access, parking, campground, 2 campground, picnic area and trails. 26.5 Knife River Marina. Access at launch area. watercraft campsites (available on a first-come, [47° 11.865' N / 91° 22.620' W] Parking, toilets. [46° 56.705' N / 91° 46.950' W] first-served basis), picnic area and trails. [47° 08.560' N / 91° 27.500' W] 59.0 Gold Rock Point. Wreck of the Madeira, driven 26.6 Knife River Beach. Carry-in access, rest area, ashore in 1905, lies scattered on the bottom in parking, toilet. Sand and pebble beach. 53.0 Thompson Beach. Four watercraft campsites 10 to 100' of water with portions clearly visible [46° 56.785' N / 91° 46.845' W] and rest area, toilet. No fires. First-come, in calm water. A popular recreational diving site, first-served. [47° 09.480' N / 91° 26.230' W] please be alert to divers in the water. Rest area 30.2 Private resort. Rocky Beach. on small beach nearby. No facilities. [46° 59.025' N / 91° 44.170' W] 53.8 Twin Points. Rest area, trailer access, parking. [47° 12.410' N / 91° 21.520' W] No camping permitted. -

Superior Hiking Trail Rises to Craggy Peaks and Plunges Into Forests of Birch, Maple, Spruce, Cedar, and Pine
Photography by Gary Alan Nelson A Trail With a View For spectacular vistas, follow a footpath along the North Shore’s rocky ridge. Are you up for a day hike in one of Minnesota’s most dramatic landscapes? The Superior Hiking Trail rises to craggy peaks and plunges into forests of birch, maple, spruce, cedar, and pine. It crosses rushing streams and opens to panoramas of Lake Superior and the highlands. Built just for hiking and backpacking, the 296-mile footpath runs from Jay Cooke State Park to the Ontario border. Each year more than 50,000 hikers explore parts of this sensational trail. With 53 trailhead parking lots, one about every 5 to 10 miles, you can easily hop on and hike for an hour or a day. Here’s a look at some of the sights along three stretches. 26 Minnesota Conservation Volunteer July–August 2014 27 Gooseberry to Split Rock Gooseberry Falls State Park is a popular starting point. In the park, a bench overlooks the Gooseberry River. Markers assure hikers they’re on trail. This 6-mile section follows Bread Loaf Ridge. Atop a cliff, hikers gain a bird’s-eye view. During spring and fall, hikers can see migratory birds along this North Shore flyway. July–August 2014 29 Waterfalls on the Gooseberry River create a soundscape. From time to time, hikers get a view of the open sky over the big lake. In the late 1890s, lumber companies logged the land along the river. By the 1920s logging and fire had cleared the pines. -

Survey and Fish Man- E Streams of the North Shore Watershed
nical Bulletin Number 1 SURVEY AND FISH MAN- E STREAMS OF THE NORTH SHORE WATERSHED LLOYD L. SM ITH, JR. and JOHN B. MOYLE DEPARTMENT Of CONSERVATION ISION OF GAME AND FISH This document is made available electronically by the Minnesota Legislative Reference Library as part of an ongoing digital archiving project. http://www.leg.state.mn.us/lrl/lrl.asp (Funding for document digitization was provided, in part, by a grant from the Minnesota Historical & Cultural Heritage Program.) MINNESOTA DEPARTMENT OF CONSERVATION DIVISION OF GAME AND FISH A BIOLOGICAL SURVEY AND FISHERY MAN AGEMENT PLAN FOR THE STREAMS OF THE LAKE SUPERIOR NORTH SHORE WATERSHED LLOYD L. SMITH, JR. Research Supervisor and JOHN B. MOYLE Aquatic Biologist A CONTRIBUTION FROM THE MINNESOTA FISHERIES RESEARCH LABORATORY TECHNICAL BULLETIN NO. 1 1 9 4 4 STATE OF MINNESOTA The Honorable Edward J. Thye ................... Governor MINNESOTA DEPARTMENT OF CONSERVATION Chester S. Wilson ............................ Commissioner E. V. Willard ........................ Deputy Commissioner DIVISION OF GAME AND FISH Verne E. Joslin ............................. Acting Director E. R. Starkweather ........................ Law Enforcement Norman L. Moe ........................... Fish Propagation George Weaver ........................ Commercial Fisheries Stoddard Robinson .................... Rough Fish Removal Lloyd L. Smith,- Jr........................ Fisheries Research Thomas Evans ........................ Stream Improvement Frank Blair .......................... ~ .. Game Management -

State of the Trails Final Report February 2017
STATE OF THE TRAILS FINAL REPORT FEBRUARY 2017 ParksCounCil of & Minn TrailsESoTA CONTENTS 3 Key Findings 4 Project Overview STATE OF THE TRAILS 6 Experiental Ratngs 7 Trail Roughness Index 8 DNR Investment Groups 10 Overview of State Trail Pavement Conditons 13 Segment-by-Segment Ratngs 13 Alex Laveau State Trail 29 Harmony Preston State Trail 14 Blazing Star State Trail 31 Heartland State Trail 15 Brown’s Creek State Trail 34 Luce Line State Trail 16 Casey Jones State Trail 36 Mill Towns State Trail 17 Central Lakes State Trail 36 Preston-Forestville State Trail 20 Cuyuna Lakes State Trail 37 Minnesota Valley State Trail 21 Douglas State Trail 38 Paul Bunyan State Trail 22 Gateway State Trail 44 Root River State Trail 23 Gitchi-Gami State Trail 47 Sakatah Singing Hills State Trail 24 Glacial Lakes State Trail 49 Shootng Star State Trail 27 Goodhue Pioneer State Trail 51 Willard Munger State Trail 28 Great River Ridge State Trail RECOMMENDATIONS 54 Consistent State Trail Rehabilitaton 57 Create a State Trail Capital Improvement Plan METHODOLOGY 59 How to Understand Our Ratngs 61 Important Caveats 63 Acknowledgments Vector artwork on cover designed by Freepik 2 www.parksandtrails.org KEY FINDINGS STATE TRAIL CONDITIONS: RECOMMENDATIONS: • Minnesota State Trails are currently in good • Lawmakers and the DNR should commit to conditon. Our inventory found 79% of state rehabilitatng an average of 20 state trail miles trail miles are in either excellent or good per year, which is the level needed to maintain conditon. Minnesota State Trails long term. • Only 26 miles of Minnesota State Trails are • Lawmakers should commit to funding state currently in poor conditon, and 90 miles are in trail rehabilitaton through consistent state fair conditon. -

Watertrail Map 2.FH10
Route Description LAKE SUPERIOR Be familiar with dangers of hypothermia and All watercraft (including non-motorized canoes and Other items recommended for paddlers to carry: (continued from other side) ake Superior is the largest freshwater dress appropriately for the cold water (32 to 50 kayaks over 9 feet in length) must be registered in A portable VHF radio to call for help in an emer- In Miles (0.0 at Minnesota Entrance -Duluth Lift Bridge) lake on our planet, containing 10% of degrees Fahrenheit). Minnesota or the state of residence. gency and monitor the weather channels; Spray skirt; Float for paddle; Whistle and emergency all the fresh water on earth. The lake's Cold water is a killer - wearing a wet or dry suit is 42.3 Crazy Bay. Split Rock Lighthouse State Park. Two 32,000 square mile surface area stretches strongly recommended. Anticipate changes in weather, wind and wave by flares; Water, snacks and sunscreen; and compass. kayak campsites. West site is for kayakers only and across the border between the United monitoring a weather or marine VHF radio, and using is available on a first-come, first-served basis. Pit States and Canada; two countries, three states, one Seek instruction and practice kayak skills, in- your awareness and common sense. This map is not adequate for sole use as a toilet. [47° 11.075' N / 91° 23.975' W]. East site (backpack/kayak site #3) is shared-use by kayakers province and many First Nations surround Superior's cluding rescues, before paddling on Lake Superior. The National Weather Service broadcasts a 24 hour navigational aid. -

ATLAS of the SPAWNING and NURSERY AREAS of GREAT LAKES FISHES Volume II - Lake Superior
Biological Services Program FWS/OBS-82/52 SEPTEMBER 1982 ATLAS OF THE SPAWNING AND NURSERY AREAS OF GREAT LAKES FISHES Volume II - Lake Superior Great Lake - St. Lawrence Seaway Navigation Season Extension Program Fish and Wildlife Service Corps of Engineers U.S. Department of the Interior U.S. Department of the Army The Biological Services Program was established within the U.S. Fish and Wildlife Service to supply scientific information and methodologies on key environmental issues that Impact fish and wildlife resources and their supporting ecosystems. The mission of the program is as follows: o To strengthen the Fish and Wildlife Service in its role as a primary source of information on national fish and wild- life resources, particularly in respect to environmental impact assessment. o To gather, analyze, and present information that will aid decisionmakers in the identification and resolution of problems associated with major changes in land and water use. o To provide better ecological information and evaluation for Department of the Interior development programs, such as those relatfng to energy development. Information developed by the Biological Services Program is intended for use in the planning and decisionmaking process to prevent or minimize the impact of development on fish and wildlife. Research activities and technlcal assistance services are based on an analysis of the issues, a determination of the decisionmakers involved and their informatlon needs, and an evaluation of the state of the art to identify information gaps and to determine priorities. This is a strategy that will ensure that the products produced and disseminated are timely and useful. -

Posted Boundaries and Fish Sanctuaries on Lake Superior Tributaries
Division of Fish and Wildlife Section of Fisheries May 2020 Posted Boundaries and Fish Sanctuaries on Lake Superior Tributaries The Minnesota Department of Natural Resources-Section of Fisheries has established regulations and fish sanctuaries on Lake Superior tributaries to protect migratory fish species from Lake Superior, particularly native coaster Brook Trout, and also to extend fishing seasons for other species. Fish sanctuaries have permanent or seasonal closures (Minnesota administrative rule 6264.0500) to protect fish in vulnerable locations during spawning seasons and to restrict fishing near dams, fish traps and egg collection stations. Fish sanctuaries are marked by signs hung by cables, attached to natural features or on posts. Posted boundaries for areas covered by Lake Superior and below-boundary tributary regulations are marked by yellow signs posted near the stream at the upstream end of the boundary. Posted boundaries specify the location on a stream where fishing regulations change and generally correspond to areas accessible to fish migrating upstream from Lake Superior. When a stream has no impassible barrier, such as a waterfall, the posted boundary is marked at a road crossing or other landmark. Streams with a posted boundary at the stream mouth or Minnesota/Wisconsin state line will not have a physical sign posted. Regulations for below posted boundary areas are: Most people fishing Lake Superior or its tributaries will need a trout/salmon stamp validation in addition to a Minnesota angling license (see MNDNR fishing regulations). Many special possession limits and size restrictions apply for trout and salmon caught below the posted boundaries (see MNDNR fishing regulations). -

Report of the Section of Mineralogy
Journal of the Minnesota Academy of Science Volume 2 Number 5 Article 5 1885 Report of the Section of Mineralogy N. H. Winchell Follow this and additional works at: https://digitalcommons.morris.umn.edu/jmas Part of the Geology Commons Recommended Citation Winchell, N. H. (1885). Report of the Section of Mineralogy. Journal of the Minnesota Academy of Science, Vol. 2 No.5, 390-416. Retrieved from https://digitalcommons.morris.umn.edu/jmas/vol2/iss5/5 This Article is brought to you for free and open access by the Journals at University of Minnesota Morris Digital Well. It has been accepted for inclusion in Journal of the Minnesota Academy of Science by an authorized editor of University of Minnesota Morris Digital Well. For more information, please contact [email protected]. REPOHT OF THE SECTIOX OF l\:!IX~~RALOGY BY N. H. Wll\('HEJ,L, ('HAIRlL\N. [Octobtor, 188:.!.} In the inception of any systematic and thorough work in the Mineralo~y of Minnesota, it is ohvious that the .tin~f thing is to find out what has been done alread~·. It is the object of this paper to accomplish that for the Academy. In other words this paper begins with a reference to publications on the min erals of the State so far as they can now be ascertained-a kind of Bibliography q( the miueralog!f qf Jlinnesota -and ends with a1~ eJtumerafiou if suclt minerals as are known to occur in the State, and a statement of their localities, with notes on some of their characters or peculiarities. -

SHTA Brochure
www.shta.org THE SUPERIOR HIKING TRAIL Information, photos, and more. Check the IS WAITING FOR YOU. “Conditions” page to get information on trail conditions. Visit the “Store” to make a donation, become a member, or shop for SHT items. THE SUPERIOR HIKING TRAIL largely follows the scenic ridgeline high above Minnesota’s Lake Superior North Shore and extends for 310 miles from Jay Cooke State Park to the We’re member supported and Canadian border. Narrow and rugged, the trail connects +,-îr /010í eight state parks, state and county forests, and the Superior National Forest. The SHT is designed for foot travel only and BECOME A MEMBER. The Superior Hiking Trail is well-marked with blue paint blazes for the main trail and Association (SHTA), a non-profit organization with white blazes for spur and overlook trails. more than 3,800 members, builds, maintains and promotes this spectacular trail. The Superior Hiking Trail depends on members for its very existence; it is not maintained by any government agency. PLAN YOUR ADVENTURE Memberships benefits include: T!e 2 "#s o 2 %&s ' ( )*l • Free trail maps DAY HIKING. Access the trail from 53 trailhead park- • Discount prices on SHTA merchandise ing lots every 3–11 miles. The trail is narrow with roots • The Ridgeline newsletter four times per year and rocks. Wear sturdy footwear and allow one hour • Monthly e-bulletin SHT News You Can Use for every one to two miles hiked. Bring water, rainwear, • SHTA events notice— guided hike schedule, annual snacks, insect repellant, and a warm layer. -

A Fishing Guide to Lake Superior and North Shore Trout Streams
AA FishingFishing GuideGuide toto LakeLake SuperiorSuperior andand NorthNorth ShoreShore TTroutrout StreamsStreams LEGEND Stream Information Seasonal Fishing Lake County Boat Launch Sites Miles Miles A. Horseshoe Bay E. Schroeder Town Launch Cook County Above Below Trout Shoreline Miles Miles (DNR) Located one and one-quarter miles east of Hovland. Turn off State Highway 61 east of Cross River on road marked Stream Name Boundary Boundary Species Status Continuous Fishing Above Below Trout Shoreline No gas. Parking. Small boats only. Father Baragas Cross, west side of Temperance River State except for brook trout Stream Name Boundary Boundary Species Status Park. The launch is just left of the dead end. No gas. Small Duluth Baptism River 8.0 1.00 B,Bn,R,C P,G B. Grand Marais boats only. Parking. Picnic area. Baptism River, E. Branch 14.0 0.00 B,Bn P,G (DNR/City) Heading north on State Highway 61 take a Assinika Creek 4.1 0.00 B G Baptism River, W. Branch 14.5 0.00 B,Bn P,G right at the stop lights in Grand Marais. Three blocks to F. Taconite Harbor Bally Creek 5.5 0.00 B G Beaver River 24.1 0.20 B,Bn,R P,G Boat Access Barker Creek 6.5 0.00 B G Beaver River, E. Branch 23.0 0.00 B P,G launch site adjacent to Coast Guard Station. No gas. Parking. (DNR) Turn at public access sign off State Highway 61 west Beaver Dam Creek 5.0 0.00 B P,G Beaver River, W. -
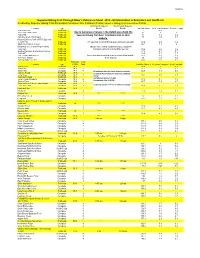
2016 SHT Reference Sheet
3/5/2016 Superior Hiking Trail Through Hiker's Reference Sheet 2016...All Information is Estimated and Unofficial Created by Superior Hiking Trail Association Volunteer Kim Fishburn © 2016, Superior Hiking Trail Association © 2016 Northbound Superior Southbound Superior Location Type Shuttle Shuttle Trail Mile Mark Next Trail Junction Section Length Wild Valley Rd Trailhead 0 5.9 Jay Cooke State Park Trailhead Due to numerous changes in the Duluth area check the 5.9 3 3 Palkie Rd Trailhead Superior Hiking Trail Assn. Conditions Tab on their 8.9 8.1 6.4 123 Ave West (150 ft spur) Trailhead 17 4.2 4.3 Magney Snively Park (350 Ft Spur trail website. to main trail) Trailhead0.5 spur trail to Spirit Mt Campground near mid point 21.2 2.9 3.2 Spirit Mountain (0.3 spur) Trailhead 24.1 1.1 2 Kingsbury Creek (600 ft spur trail to Munger Inn located southwest of zoo, Indian Pt main trail) TrailheadCampground located behind Munger Inn 25.2 2.6 3.2 Skyline Parkway at Highland/Getchell Trailhead 27.8 5.7 5.7 th North 24 Trailhead 33.5 1.7 1.7 Enger Park (440 ft spur) Trailhead Numerous hotels located beween Enger Park and the 35.2 3.8 3.8 The Rose Garden Trailhead Rose Garden 39 4.5 4.5 Hartley Nature Center Trailhead 43.5 3.1 3.1 Highway Tent Location Type 61 Mile Pads Trail Mile Marker Previous Campsite Next Campsite Martin Road Trailhead 14.9 **** **** 46.6 0 4.9 Bald Eagle Campsite 14.9 3 51.5 0 1.1 White Pine Campsite 14.9 4 All trailheads from Jay Cooke State Park to Lake 52.6 1.1 5.5 Lismore Road Trailhead 14.9 Co.