Final Report: JFSP Project Number 14-3-01-30
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Box Turtles July 2017
The HERP Project, Herpetology Education in Rural Places and Spaces In Awe of Nature: Treasuring Terrestrial Turtles By Ann Berry Somers, Catherine Matthews, and Lacey Huffling The Herp Project is supported by the National Science Foundation, Grant No. DRL-1114558. Any opinions, findings, and conclusions or recommendations expressed in this manuscript are those of the authors and do not necessarily reflect the views of the National Science Foundation. Treasuring Terrestrial Turtles Before starting a project like the one described in this curriculum, contact your state wildlife resources commission or state division of fish and game to see what permits you need to work with box turtles. I. Project Description This curriculum was developed by The HERP (Herpetology Education in Rural Places and Spaces) Project to introduce participants to the wonders of nature and science through the study of box turtles (Terrapene spp.). The curriculum was developed over several years of working with high school students in our Herpetological Research Experience (HRE) residential program. Feel free to modify this curriculum as needed. In our program, participants are introduced to turtle biology as well as to The Box Turtle Connection (BTC), our long-term mark/recapture box turtle study in North Carolina. The BTC is designed to follow temporal trends in population size and structure (sex, age class) as well as the health and condition of individual box turtles from numerous sites across North Carolina. The data collected are important to help scientists determine if box turtles need special conservation measures to maintain their populations and thrive in their natural habitat. Our box turtle studies are enhanced by use of Boykin Spaniel dogs to locate and retrieve box turtles and use of radio tracking to determine activity ranges for male and female box turtles. -

AN INTRODUCTION to Texas Turtles
TEXAS PARKS AND WILDLIFE AN INTRODUCTION TO Texas Turtles Mark Klym An Introduction to Texas Turtles Turtle, tortoise or terrapin? Many people get confused by these terms, often using them interchangeably. Texas has a single species of tortoise, the Texas tortoise (Gopherus berlanderi) and a single species of terrapin, the diamondback terrapin (Malaclemys terrapin). All of the remaining 28 species of the order Testudines found in Texas are called “turtles,” although some like the box turtles (Terrapene spp.) are highly terrestrial others are found only in marine (saltwater) settings. In some countries such as Great Britain or Australia, these terms are very specific and relate to the habit or habitat of the animal; in North America they are denoted using these definitions. Turtle: an aquatic or semi-aquatic animal with webbed feet. Tortoise: a terrestrial animal with clubbed feet, domed shell and generally inhabiting warmer regions. Whatever we call them, these animals are a unique tie to a period of earth’s history all but lost in the living world. Turtles are some of the oldest reptilian species on the earth, virtually unchanged in 200 million years or more! These slow-moving, tooth less, egg-laying creatures date back to the dinosaurs and still retain traits they used An Introduction to Texas Turtles | 1 to survive then. Although many turtles spend most of their lives in water, they are air-breathing animals and must come to the surface to breathe. If they spend all this time in water, why do we see them on logs, rocks and the shoreline so often? Unlike birds and mammals, turtles are ectothermic, or cold- blooded, meaning they rely on the temperature around them to regulate their body temperature. -
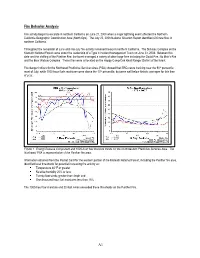
A1 Fire Behavior Analysis
Fire Behavior Analysis Fire activity began to escalate in northern California on June 21, 2008 when a major lightning event affected the Northern California Geographic Coordination Area (North Ops). The July 22, 2008 National Situation Report identified 602 new fires in northern California. Throughout the remainder of June and into July fire activity remained heavy in northern California. The Siskiyou Complex on the Klamath National Forest came under the leadership of a Type II Incident Management Team on June 23, 2008. Between this date and the staffing of the Panther Fire, the forest managed a variety of other large fires including the Gould Fire, No Man‘s Fire and the Bear Wallow Complex. These fires were all located on the Happy Camp/Oak Knoll Ranger District of the forest. Fire danger indices for the Northwest Predictive Services Area (PSA) showed that ERCs were tracking near the 90th percentile most of July, while 1000-hour fuels moistures were above the 10th percentile, but were well below historic averages for this time of year. Figure 1. Energy Release Component and 1000-hour fuel moisture trends for the Northwestern Predictive Services Area. The Northwest PSA is representative of the Panther fire area. Information obtained from the Pocket Card for the western portion of the Klamath National Forest, including the Panther fire area, identified local thresholds for potential increasing fire activity as: Temperature 85°F or greater Relative humidity 20% or less Twenty-foot winds greater than 4mph and One-thousand hour fuel moistures less than 15% The 1000-hour fuel moisture and 20-foot winds exceeded these thresholds on the Panther Fire. -

In AR, FL, GA, IA, KY, LA, MO, OH, OK, SC, TN, and TX): Species in Red = Depleted to the Point They May Warrant Federal Endangered Species Act Listing
Southern and Midwestern Turtle Species Affected by Commercial Harvest (in AR, FL, GA, IA, KY, LA, MO, OH, OK, SC, TN, and TX): species in red = depleted to the point they may warrant federal Endangered Species Act listing Common snapping turtle (Chelydra serpentina) – AR, GA, IA, KY, MO, OH, OK, SC, TX Florida common snapping turtle (Chelydra serpentina osceola) - FL Southern painted turtle (Chrysemys dorsalis) – AR Western painted turtle (Chrysemys picta) – IA, MO, OH, OK Spotted turtle (Clemmys gutatta) - FL, GA, OH Florida chicken turtle (Deirochelys reticularia chrysea) – FL Western chicken turtle (Deirochelys reticularia miaria) – AR, FL, GA, KY, MO, OK, TN, TX Barbour’s map turtle (Graptemys barbouri) - FL, GA Cagle’s map turtle (Graptemys caglei) - TX Escambia map turtle (Graptemys ernsti) – FL Common map turtle (Graptemys geographica) – AR, GA, OH, OK Ouachita map turtle (Graptemys ouachitensis) – AR, GA, OH, OK, TX Sabine map turtle (Graptemys ouachitensis sabinensis) – TX False map turtle (Graptemys pseudogeographica) – MO, OK, TX Mississippi map turtle (Graptemys pseuogeographica kohnii) – AR, TX Alabama map turtle (Graptemys pulchra) – GA Texas map turtle (Graptemys versa) - TX Striped mud turtle (Kinosternon baurii) – FL, GA, SC Yellow mud turtle (Kinosternon flavescens) – OK, TX Common mud turtle (Kinosternon subrubrum) – AR, FL, GA, OK, TX Alligator snapping turtle (Macrochelys temminckii) – AR, FL, GA, LA, MO, TX Diamond-back terrapin (Malaclemys terrapin) – FL, GA, LA, SC, TX River cooter (Pseudemys concinna) – AR, FL, -

Introduction to Fire Behavior Modeling (2012)
Introduction to Fire Behavior Modeling Introduction to Wildfire Behavior Modeling Introduction Table of Contents Introduction ........................................................................................................ 5 Chapter 1: Background........................................................................................ 7 What is wildfire? ..................................................................................................................... 7 Wildfire morphology ............................................................................................................. 10 By shape........................................................................................................ 10 By relative spread direction ........................................................................... 12 Wildfire behavior characteristics ........................................................................................... 14 Flame front rate of spread (ROS) ................................................................... 15 Heat per unit area (HPA) ................................................................................ 17 Fireline intensity (FLI) .................................................................................... 19 Flame size ..................................................................................................... 23 Major influences on fire behavior simulations ....................................................................... 24 Fuelbed structure ......................................................................................... -

Habitat Alteration and Survival Rates of the Ornate Box Turtle Sarah M
Ecology, Evolution and Organismal Biology Ecology, Evolution and Organismal Biology Publications 11-2016 Habitat alteration and survival rates of the ornate box turtle Sarah M. Mitchell Iowa State University Brooke L. Bodensteiner Iowa State University Jeramie T. Strickland U.S. Fish and Wildlife Service James K. Quick North Carolina A & T State University Fredric J. Janzen Iowa State University, [email protected] Follow this and additional works at: https://lib.dr.iastate.edu/eeob_ag_pubs Part of the Animal Sciences Commons, and the Terrestrial and Aquatic Ecology Commons The ompc lete bibliographic information for this item can be found at https://lib.dr.iastate.edu/ eeob_ag_pubs/275. For information on how to cite this item, please visit http://lib.dr.iastate.edu/ howtocite.html. This Article is brought to you for free and open access by the Ecology, Evolution and Organismal Biology at Iowa State University Digital Repository. It has been accepted for inclusion in Ecology, Evolution and Organismal Biology Publications by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. Habitat alteration and survival rates of the ornate box turtle Abstract Habitat destruction and modification may be the most prominent anthropogenic forces affecting extant biological systems. Growing evidence suggests that turtles are especially vulnerable to many anthropogenic stressors. We evaluated the effects of habitat modification on survival rates of the threatened ornate box turtle (Terrapene ornata) in northwest Illinois, USA, using a 20‐year mark‐recapture dataset. Longstanding development (i.e., cottages, outbuildings, landscape management) reduced the apparent survival of the ornate box turtle, especially among females. -

Box Turtle Care Compiled by Dayna Willems, DVM
Box Turtle Care Compiled by Dayna Willems, DVM Brief Description There are several species of box turtle native to North America (ornate box turtles in Colorado) but their wild populations are declining rapidly due to habitat loss, being hit by cars on roads, and from collection for the pet trade. Unfortunately once taken from the wild box turtles can have trouble adapting to captivity, however unless they can be released in the location they were found they will wander aimlessly searching for their lost territory for months often into roads or neighborhoods. The box turtle is known for being able to completely withdraw its body into its protective shell and a hinge on the bottom part of the shell allows it to close completely like a box. The shell is living tissue and should never be pierced or painted. Three-toed Box Turtle Lifespan With good care the expected lifespan is 60-80 years on average. Sexing Once mature male box turtles will usually have red eyes and females will have brown eyes in most cases, however not always. Males will have an indent on their plastron (belly of the shell) and a longer, more pointed tail with a longer distance between vent and tail tip than the stubby tail of females where the vent is closer to the shell. Ornate Box Turtle Caging Box turtles need large enclosures and when able should be housed outdoors in a protected outdoor pen. Box turtles are great climbers so the pen will need to be secure to keep turtles in and predators (especially dogs) out. -

8123 Songs, 21 Days, 63.83 GB
Page 1 of 247 Music 8123 songs, 21 days, 63.83 GB Name Artist The A Team Ed Sheeran A-List (Radio Edit) XMIXR Sisqo feat. Waka Flocka Flame A.D.I.D.A.S. (Clean Edit) Killer Mike ft Big Boi Aaroma (Bonus Version) Pru About A Girl The Academy Is... About The Money (Radio Edit) XMIXR T.I. feat. Young Thug About The Money (Remix) (Radio Edit) XMIXR T.I. feat. Young Thug, Lil Wayne & Jeezy About Us [Pop Edit] Brooke Hogan ft. Paul Wall Absolute Zero (Radio Edit) XMIXR Stone Sour Absolutely (Story Of A Girl) Ninedays Absolution Calling (Radio Edit) XMIXR Incubus Acapella Karmin Acapella Kelis Acapella (Radio Edit) XMIXR Karmin Accidentally in Love Counting Crows According To You (Top 40 Edit) Orianthi Act Right (Promo Only Clean Edit) Yo Gotti Feat. Young Jeezy & YG Act Right (Radio Edit) XMIXR Yo Gotti ft Jeezy & YG Actin Crazy (Radio Edit) XMIXR Action Bronson Actin' Up (Clean) Wale & Meek Mill f./French Montana Actin' Up (Radio Edit) XMIXR Wale & Meek Mill ft French Montana Action Man Hafdís Huld Addicted Ace Young Addicted Enrique Iglsias Addicted Saving abel Addicted Simple Plan Addicted To Bass Puretone Addicted To Pain (Radio Edit) XMIXR Alter Bridge Addicted To You (Radio Edit) XMIXR Avicii Addiction Ryan Leslie Feat. Cassie & Fabolous Music Page 2 of 247 Name Artist Addresses (Radio Edit) XMIXR T.I. Adore You (Radio Edit) XMIXR Miley Cyrus Adorn Miguel Adorn Miguel Adorn (Radio Edit) XMIXR Miguel Adorn (Remix) Miguel f./Wiz Khalifa Adorn (Remix) (Radio Edit) XMIXR Miguel ft Wiz Khalifa Adrenaline (Radio Edit) XMIXR Shinedown Adrienne Calling, The Adult Swim (Radio Edit) XMIXR DJ Spinking feat. -

English Song Booklet
English Song Booklet SONG NUMBER SONG TITLE SINGER SONG NUMBER SONG TITLE SINGER 100002 1 & 1 BEYONCE 100003 10 SECONDS JAZMINE SULLIVAN 100007 18 INCHES LAUREN ALAINA 100008 19 AND CRAZY BOMSHEL 100012 2 IN THE MORNING 100013 2 REASONS TREY SONGZ,TI 100014 2 UNLIMITED NO LIMIT 100015 2012 IT AIN'T THE END JAY SEAN,NICKI MINAJ 100017 2012PRADA ENGLISH DJ 100018 21 GUNS GREEN DAY 100019 21 QUESTIONS 5 CENT 100021 21ST CENTURY BREAKDOWN GREEN DAY 100022 21ST CENTURY GIRL WILLOW SMITH 100023 22 (ORIGINAL) TAYLOR SWIFT 100027 25 MINUTES 100028 2PAC CALIFORNIA LOVE 100030 3 WAY LADY GAGA 100031 365 DAYS ZZ WARD 100033 3AM MATCHBOX 2 100035 4 MINUTES MADONNA,JUSTIN TIMBERLAKE 100034 4 MINUTES(LIVE) MADONNA 100036 4 MY TOWN LIL WAYNE,DRAKE 100037 40 DAYS BLESSTHEFALL 100038 455 ROCKET KATHY MATTEA 100039 4EVER THE VERONICAS 100040 4H55 (REMIX) LYNDA TRANG DAI 100043 4TH OF JULY KELIS 100042 4TH OF JULY BRIAN MCKNIGHT 100041 4TH OF JULY FIREWORKS KELIS 100044 5 O'CLOCK T PAIN 100046 50 WAYS TO SAY GOODBYE TRAIN 100045 50 WAYS TO SAY GOODBYE TRAIN 100047 6 FOOT 7 FOOT LIL WAYNE 100048 7 DAYS CRAIG DAVID 100049 7 THINGS MILEY CYRUS 100050 9 PIECE RICK ROSS,LIL WAYNE 100051 93 MILLION MILES JASON MRAZ 100052 A BABY CHANGES EVERYTHING FAITH HILL 100053 A BEAUTIFUL LIE 3 SECONDS TO MARS 100054 A DIFFERENT CORNER GEORGE MICHAEL 100055 A DIFFERENT SIDE OF ME ALLSTAR WEEKEND 100056 A FACE LIKE THAT PET SHOP BOYS 100057 A HOLLY JOLLY CHRISTMAS LADY ANTEBELLUM 500164 A KIND OF HUSH HERMAN'S HERMITS 500165 A KISS IS A TERRIBLE THING (TO WASTE) MEAT LOAF 500166 A KISS TO BUILD A DREAM ON LOUIS ARMSTRONG 100058 A KISS WITH A FIST FLORENCE 100059 A LIGHT THAT NEVER COMES LINKIN PARK 500167 A LITTLE BIT LONGER JONAS BROTHERS 500168 A LITTLE BIT ME, A LITTLE BIT YOU THE MONKEES 500170 A LITTLE BIT MORE DR. -

A Field Guide to South Dakota Turtles
A Field Guide to SOUTH DAKOTA TURTLES EC919 South Dakota State University | Cooperative Extension Service | USDA U.S. Geological Survey | South Dakota Cooperative Fish and Wildlife Research Unit South Dakota Department of Game, Fish & Parks This publication may be cited as: Bandas, Sarah J., and Kenneth F. Higgins. 2004. Field Guide to South Dakota Turtles. SDCES EC 919. Brookings: South Dakota State University. Copies may be obtained from: Dept. of Wildlife & Fisheries Sciences South Dakota State University Box 2140B, NPBL Brookings SD 57007-1696 South Dakota Dept of Game, Fish & Parks 523 E. Capitol, Foss Bldg Pierre SD 57501 SDSU Bulletin Room ACC Box 2231 Brookings, SD 57007 (605) 688–4187 A Field Guide to SOUTH DAKOTA TURTLES EC919 South Dakota State University | Cooperative Extension Service | USDA U.S. Geological Survey | South Dakota Cooperative Fish and Wildlife Research Unit South Dakota Department of Game, Fish & Parks Sarah J. Bandas Department of Wildlife and Fisheries Sciences South Dakota State University NPB Box 2140B Brookings, SD 57007 Kenneth F. Higgins U.S. Geological Survey South Dakota Cooperative Fish and Wildlife Research Unit South Dakota State University NPB Box 2140B Brookings, SD 57007 Contents 2 Introduction . .3 Status of South Dakota turtles . .3 Fossil record and evolution . .4 General turtle information . .4 Taxonomy of South Dakota turtles . .9 Capturing techniques . .10 Turtle handling . .10 Turtle habitats . .13 Western Painted Turtle (Chrysemys picta bellii) . .15 Snapping Turtle (Chelydra serpentina) . .17 Spiny Softshell Turtle (Apalone spinifera) . .19 Smooth Softshell Turtle (Apalone mutica) . .23 False Map Turtle (Graptemys pseudogeographica) . .25 Western Ornate Box Turtle (Terrapene ornata ornata) . -

Frequently Asked Questions on Flame Retardants
Flame Retardants Frequently Asked Questions The European Flame Retardants Association EFRA - The European Flame Retardants Association Chemistry making a world of difference Compiled and edited by Adrian Beard, Clariant, fire test photos by Ralf Baumgarten taken at Siemens Brandversuchshaus in Frankfurt- Hoechst and BayerIndustryServices Fire Test Laboratory in Leverkusen. We thank the teams of Knut Bauer and Michael Halfmann for their help and technical assistance. Cover photos: Candles are a common cause of ignition for domestic fires. The stereo shown on the front cover is engulfed in flames after 7 minutes when ignited with a small flame - see the photo on the back cover. Contents Flame Retardants Frequently Asked Questions Flame Retardants - General Aspects 2 General Fire Safety 4 Fire Safety Standards and Regulations 8 Flame Retardants and other Safety Technologies 12 Flame Retardant types and applications 14 Brominated Flame Retardants (BFRs) 16 Flame Retardants based on Phosphorus Compounds (PFRs) 18 Mineral flame retardants 20 Nitrogen-containing Flame Retardants 22 Other Flame Retardants - Borates, Stannates, ... 24 Flame Retardants - Health and the Environment 26 Recycling and Waste Management of Flame Retardants 32 Common Abbreviations for Flame Retardants 35 Literature and Further Reading 36 EFRA Members 37 1 Flame Retardants Frequently asked Questions Flame Retardants - General Aspects What are flame retardants? Flame retardants are chemicals which are added to combustible materials to render them more resistant to ignition. They are designed to minimise the risk of a fire starting in case of contact with a small heat source such as a cigarette, candle or an electrical fault. If the flame retarded material or an adjacent material has ignited, the flame retardant will slow down combustion and often prevent the fire from spreading to other items. -

Boiler Manual • Installation • Maintenance • Startup • Parts
Gas-Fired Water Boilers WM97+70/110/155 - CT Boiler Manual • Installation • Maintenance • Startup • Parts Now available Matching High Performance Companion Water Heater (Unit sold separately) This manual must only be used by a qualified heating installer/service technician. Read all instructions, includ- ing this manual and all other information shipped with the boiler, before installing. Perform steps in the order given. Failure to comply could result in severe personal injury, death or substantial property damage. Part number 550-142-120/0113 WM97+ GAS-FIRED WATER BOILER — Boiler Manual The WM97+70 & 110 Gas-fired water boilers 1. Stainless steel firetube heat exchanger to match boiler output to space heating and/or DHW heating demand. The control provides for three heating zones (DHW plus 2. Heat exchanger access cover/burner mounting plate two space-heating or three space heating). DHW direct-connected 3. Blower to the bottom piping connections can be set for domestic priority. The advanced blower design and air inlet silencer on WM97+ boilers 17. Multi-position WM97+ control housing result in very quiet operation. The WM97+ control is mounted in a plastic housing assembly that Air enters the boiler enclosure through the air intake adapter (18), can rotate forward for rear access to the control boards or for access flows through the enclosure, enters the air inlet silencer (5), then to boiler components. It can also be removed through slots in the enters the blower. The blower pulls air through the silencer and boiler jacket sides. pushes it into the venturi (6), where it mixes with gas before enter- ing the burner.