Hakea Leaf Shape
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
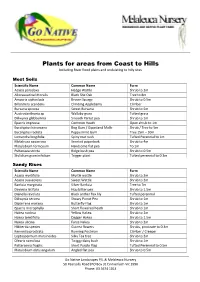
Plants for Areas from Coast to Hills Including River Flood Plains and Undulating to Hilly Sites
Plants for areas from Coast to Hills Including River flood plains and undulating to hilly sites Most Soils Scientific Name Common Name Form Acacia paradoxa Hedge Wattle Shrub to 2m Allocasuarina littoralis Black She Oak Tree to 8m Amperia xiphoclada Broom Spurge Shrub to 0.5m Billardiera scandens Climbing Appleberry Climber Bursaria spinosa Sweet Bursaria Shrub to 5m Austrodanthonia sp Wallaby grass Tufted grass Dillwynia glabberima Smooth Parrot pea Shrub to 1m Epacris impressa Common Heath Open shrub to 1m Eucalyptus kitsoniana Bog Gum / Gippsland Malle Shrub / Tree to 5m Eucalyptus radiata Peppermint Gum Tree 15m – 30m Lomandra longifolia Spiny mat rush Tufted Perennial to 1m Melaleuca squarrosa Scented paperbark Shrub to 4m Platylobium formosum Handsome flat pea To 1m Pultenaea stricta Ridge bush pea Shrub to 0.5m Stylidium graminifolium Trigger plant Tufted perennial to 0.3m Sandy Rises Scientific Name Common Name Form Acacia myrtifolia Myrtle wattle Shrub to 2m Acacia suaveolens Sweet Wattle Shrub to 2m Banksia marginata Silver Banksia Tree to 7m Daviesia latifolia Hop bitter pea Shrub to 1.5m Dianella revoluta Black anther flax lily Tufted perennial Dillwynia sericea Showy Parrot Pea Shrub to 1m Diplarrena moraea Butterfly Flag Shrub to 1m Epacris microphylla Short flowered heath Shrub to 1m Hakea nodosa Yellow Hakea Shrub to 2m Hakea teretifolia Dagger Hakea Shrub to 1.5m Hakea ulicina Furze Hakea Shrub to 2m Hibbertia species Guinea flowers Shrubs, prostrate to 0.3m Kennedia prostrata Running Postman Climber / Creeper Leptospermum -

PUBLISHER S Candolle Herbarium
Guide ERBARIUM H Candolle Herbarium Pamela Burns-Balogh ANDOLLE C Jardin Botanique, Geneva AIDC PUBLISHERP U R L 1 5H E R S S BRILLB RI LL Candolle Herbarium Jardin Botanique, Geneva Pamela Burns-Balogh Guide to the microform collection IDC number 800/2 M IDC1993 Compiler's Note The microfiche address, e.g. 120/13, refers to the fiche number and secondly to the individual photograph on each fiche arranged from left to right and from the top to the bottom row. Pamela Burns-Balogh Publisher's Note The microfiche publication of the Candolle Herbarium serves a dual purpose: the unique original plants are preserved for the future, and copies can be made available easily and cheaply for distribution to scholars and scientific institutes all over the world. The complete collection is available on 2842 microfiche (positive silver halide). The order number is 800/2. For prices of the complete collection or individual parts, please write to IDC Microform Publishers, P.O. Box 11205, 2301 EE Leiden, The Netherlands. THE DECANDOLLEPRODROMI HERBARIUM ALPHABETICAL INDEX Taxon Fiche Taxon Fiche Number Number -A- Acacia floribunda 421/2-3 Acacia glauca 424/14-15 Abatia sp. 213/18 Acacia guadalupensis 423/23 Abelia triflora 679/4 Acacia guianensis 422/5 Ablania guianensis 218/5 Acacia guilandinae 424/4 Abronia arenaria 2215/6-7 Acacia gummifera 421/15 Abroniamellifera 2215/5 Acacia haematomma 421/23 Abronia umbellata 221.5/3-4 Acacia haematoxylon 423/11 Abrotanella emarginata 1035/2 Acaciahastulata 418/5 Abrus precatorius 403/14 Acacia hebeclada 423/2-3 Acacia abietina 420/16 Acacia heterophylla 419/17-19 Acacia acanthocarpa 423/16-17 Acaciahispidissima 421/22 Acacia alata 418/3 Acacia hispidula 419/2 Acacia albida 422/17 Acacia horrida 422/18-20 Acacia amara 425/11 Acacia in....? 423/24 Acacia amoena 419/20 Acacia intertexta 421/9 Acacia anceps 419/5 Acacia julibross. -

N E W S L E T T E R
N E W S L E T T E R PLANTS OF TASMANIA Nursery and Gardens 65 Hall St Ridgeway TAS 7054 Open 7 Days a week – 9 am to 5 pm Closed Christmas Day, Boxing Day and Good Friday Phone: (03) 6239 1583 Fax: (03) 6239 1106 Email: [email protected] Newsletter 26 Spring 2011 Website: www.potn.com.au Hello, and welcome to the spring newsletter for 2011! News from the Nursery We are madly propagating at the moment, with many thousands of new cuttings putting their roots out and seedlings popping their heads up above the propagating mix. It is always an exciting time, as we experiment with seed from new species – sometimes they work, and sometimes we understand why we’ve never grown them before... New plants should start being put out into the sales area soon – fresh-faced little things ready to pop into the ground! We have recently purchased a further block of land from the ex-neighbours Jubilee Nursery, now sadly closed, that will give us a lot more flexibility and the ability to grow and store more plants. As mentioned last newsletter we have done some major revamping in the garden. A lot of work by all the staff has led to a much more open garden with a lovely Westringia brevifolia hedge (well, it will be a hedge when it grows a bit), another Micrantheum hexandrum Cream Cascade hedge-to-be, lots of Correas, Lomatias and Baueras. Where we sell a few forms of a particular species we have tried to plant examples of each so that we can show you what they are like. -

Museum of Economic Botany, Kew. Specimens Distributed 1901 - 1990
Museum of Economic Botany, Kew. Specimens distributed 1901 - 1990 Page 1 - https://biodiversitylibrary.org/page/57407494 15 July 1901 Dr T Johnson FLS, Science and Art Museum, Dublin Two cases containing the following:- Ackd 20.7.01 1. Wood of Chloroxylon swietenia, Godaveri (2 pieces) Paris Exibition 1900 2. Wood of Chloroxylon swietenia, Godaveri (2 pieces) Paris Exibition 1900 3. Wood of Melia indica, Anantapur, Paris Exhibition 1900 4. Wood of Anogeissus acuminata, Ganjam, Paris Exhibition 1900 5. Wood of Xylia dolabriformis, Godaveri, Paris Exhibition 1900 6. Wood of Pterocarpus Marsupium, Kistna, Paris Exhibition 1900 7. Wood of Lagerstremia parviflora, Godaveri, Paris Exhibition 1900 8. Wood of Anogeissus latifolia , Godaveri, Paris Exhibition 1900 9. Wood of Gyrocarpus jacquini, Kistna, Paris Exhibition 1900 10. Wood of Acrocarpus fraxinifolium, Nilgiris, Paris Exhibition 1900 11. Wood of Ulmus integrifolia, Nilgiris, Paris Exhibition 1900 12. Wood of Phyllanthus emblica, Assam, Paris Exhibition 1900 13. Wood of Adina cordifolia, Godaveri, Paris Exhibition 1900 14. Wood of Melia indica, Anantapur, Paris Exhibition 1900 15. Wood of Cedrela toona, Nilgiris, Paris Exhibition 1900 16. Wood of Premna bengalensis, Assam, Paris Exhibition 1900 17. Wood of Artocarpus chaplasha, Assam, Paris Exhibition 1900 18. Wood of Artocarpus integrifolia, Nilgiris, Paris Exhibition 1900 19. Wood of Ulmus wallichiana, N. India, Paris Exhibition 1900 20. Wood of Diospyros kurzii , India, Paris Exhibition 1900 21. Wood of Hardwickia binata, Kistna, Paris Exhibition 1900 22. Flowers of Heterotheca inuloides, Mexico, Paris Exhibition 1900 23. Leaves of Datura Stramonium, Paris Exhibition 1900 24. Plant of Mentha viridis, Paris Exhibition 1900 25. Plant of Monsonia ovata, S. -

Australian Native Plants Society Australia Hakea
AUSTRALIAN NATIVE PLANTS SOCIETY AUSTRALIA HAKEA STUDY GROUP NEWSLETTER No. 65 OCTOBER 2017 ISSN0727- 7008 Leader: Paul Kennedy 210 Aireys Street Elliminyt Vic. 3250 E mail [email protected] Tel. 03-52315569 Dear members. We have had a very cold winter and now as spring emerges the cold remains with very wet conditions. Oh how I long for some warm sunshine to brighten our day. However the Hakeas have stood up to the cold weather very well and many have now flowered. Rainfall in August was 30mm but in the first 6 days of September another 56mm was recorded making the soil very moist indeed. The rain kept falling in September with 150mm recorded. Fortunately my drainage work of spoon drains and deeper drains with slotted pipe with blue metal cover on top shed a lot of water straight into the Council drains. Most of the Hakeas like well drained conditions, so building up beds and getting rid of excess water will help in making them survive. The collection here now stands at 162 species out of a possible 169. Seed of some of the remaining species hopefully will arrive here before Christmas so that I can propagate them over summer. Wanderings. Barbara and I spent most of June and July in northern NSW and Queensland to escape the cold conditions down here. I did look around for Hakeas and visited some members’ gardens. Just to the east of Cann River I found Hakea decurrens ssp. physocarpa, Hakea ulicina and Hakea teretifolia ssp. hirsuta all growing on the edge of a swamp. -

East Gippsland, Victoria
Biodiversity Summary for NRM Regions Species List What is the summary for and where does it come from? This list has been produced by the Department of Sustainability, Environment, Water, Population and Communities (SEWPC) for the Natural Resource Management Spatial Information System. The list was produced using the AustralianAustralian Natural Natural Heritage Heritage Assessment Assessment Tool Tool (ANHAT), which analyses data from a range of plant and animal surveys and collections from across Australia to automatically generate a report for each NRM region. Data sources (Appendix 2) include national and state herbaria, museums, state governments, CSIRO, Birds Australia and a range of surveys conducted by or for DEWHA. For each family of plant and animal covered by ANHAT (Appendix 1), this document gives the number of species in the country and how many of them are found in the region. It also identifies species listed as Vulnerable, Critically Endangered, Endangered or Conservation Dependent under the EPBC Act. A biodiversity summary for this region is also available. For more information please see: www.environment.gov.au/heritage/anhat/index.html Limitations • ANHAT currently contains information on the distribution of over 30,000 Australian taxa. This includes all mammals, birds, reptiles, frogs and fish, 137 families of vascular plants (over 15,000 species) and a range of invertebrate groups. Groups notnot yet yet covered covered in inANHAT ANHAT are notnot included included in in the the list. list. • The data used come from authoritative sources, but they are not perfect. All species names have been confirmed as valid species names, but it is not possible to confirm all species locations. -

Phytophthora Resistance and Susceptibility Stock List
Currently known status of the following plants to Phytophthora species - pathogenic water moulds from the Agricultural Pathology & Kingdom Protista. Biological Farming Service C ompiled by Dr Mary Cole, Agpath P/L. Agricultural Consultants since 1980 S=susceptible; MS=moderately susceptible; T= tolerant; MT=moderately tolerant; ?=no information available. Phytophthora status Life Form Botanical Name Family Common Name Susceptible (S) Tolerant (T) Unknown (UnK) Shrub Acacia brownii Mimosaceae Heath Wattle MS Tree Acacia dealbata Mimosaceae Silver Wattle T Shrub Acacia genistifolia Mimosaceae Spreading Wattle MS Tree Acacia implexa Mimosaceae Lightwood MT Tree Acacia leprosa Mimosaceae Cinnamon Wattle ? Tree Acacia mearnsii Mimosaceae Black Wattle MS Tree Acacia melanoxylon Mimosaceae Blackwood MT Tree Acacia mucronata Mimosaceae Narrow Leaf Wattle S Tree Acacia myrtifolia Mimosaceae Myrtle Wattle S Shrub Acacia myrtifolia Mimosaceae Myrtle Wattle S Tree Acacia obliquinervia Mimosaceae Mountain Hickory Wattle ? Shrub Acacia oxycedrus Mimosaceae Spike Wattle S Shrub Acacia paradoxa Mimosaceae Hedge Wattle MT Tree Acacia pycnantha Mimosaceae Golden Wattle S Shrub Acacia sophorae Mimosaceae Coast Wattle S Shrub Acacia stricta Mimosaceae Hop Wattle ? Shrubs Acacia suaveolens Mimosaceae Sweet Wattle S Tree Acacia ulicifolia Mimosaceae Juniper Wattle S Shrub Acacia verniciflua Mimosaceae Varnish wattle S Shrub Acacia verticillata Mimosaceae Prickly Moses ? Groundcover Acaena novae-zelandiae Rosaceae Bidgee-Widgee T Tree Allocasuarina littoralis Casuarinaceae Black Sheoke S Tree Allocasuarina paludosa Casuarinaceae Swamp Sheoke S Tree Allocasuarina verticillata Casuarinaceae Drooping Sheoak S Sedge Amperea xipchoclada Euphorbaceae Broom Spurge S Grass Amphibromus neesii Poaceae Swamp Wallaby Grass ? Shrub Aotus ericoides Papillionaceae Common Aotus S Groundcover Apium prostratum Apiaceae Sea Celery MS Herb Arthropodium milleflorum Asparagaceae Pale Vanilla Lily S? Herb Arthropodium strictum Asparagaceae Chocolate Lily S? Shrub Atriplex paludosa ssp. -

INDIGENOUS Tube Stocklist 2019 Plants from the Greater Melbourne Area
INDIGENOUS tube stocklist 2019 plants from the greater Melbourne area Kuranga Native Nursery grows an extensive If you are requiring a species not listed, please range of plants which occur naturally in the enquire as it may still be available. Greater Melbourne area. Bushfood plants are We have a range of books for sale which list marked # (refer to details on back page). the indigenous plants of various areas around All plants on this list are grown in 50 mm (2”) Melbourne. square forestry tubes. The seed and All tubes are $2.95 each except those marked cutting material for most of our tube stock with * which are $3.95 each has originated from the Melbourne metropolitan area. 118 York Rd Mt Evelyn Victoria 3796 Tel:9760 8100 www.kuranga.com.au All tubes are $2.95 except those marked with * which are $3.95 each. Total price including GST. # Bushfood plants Bird attracting plants AQUATICS AND BOG PLANTS (located near pond) Baloskion tetraphyllum* Tassel Cord-rush Lovely clump, decorative foliage, reddish/brown tassels Sp-S. Shallow water. Carex appressa Tall Sedge Tussock. 40-60cm x 40-60cm. Yellow/brownish spikelets Sp-S. Moist-wet. Carex breviculmis Short-stem Sedge Tufted sedge 15cm high. Flr spikes Sp-S. Moist depressions to exposed slopes. Carex fascicularis Tassel Sedge Drooping spikelets Sp-S. 1m x 1m. Sun or shade. Pond or bog plant. Carex gaudichaudiana* Tufted Sedge Tufted sedge.10-90cm high. Erect flr spikes Sp-S. Sun, semi-shade. Water’s edge, bog. Carex tasmanica Curly Sedge Tufted sedge to 40cm high. -

Natural Versus Supplemental Diets in Possums
Natural versus Supplemental diets in possums Michelle Thomas Animalia Wildlife Shelter [email protected] 0435822699 When I first started caring for wildlife 20 years ago I started with possums. I made every possible mistake, many of which I attributed to deficient diet and lack of knowledge. My first advice on what possums ate came from a zoo who told me to feed fruit & vegetables, peppermint gum and bottlebrush. This did not seem logical, however, and I puzzled over why we should feed fruit and vegetables to native animals when such foods were not available to them in the wild or their natural habitat across the country. I bumbled along offering rose petals, geraniums, oak , willow and liquid amber leaf , none of it remotely native. By this stage the brushtail and ringtail possums in my care had been tried and tested on broccoli, cauliflower, peas, capsicum, apple, pear, watermelon, cantaloupe, sweet potato, beans, cucumber, cooked carrot, pasta, pine nuts, sunflower seeds, pumpkin seeds, peanut butter, button mushroom, banana, assorted stone fruits, peanuts, almonds, pine nuts, cashews, figs, cooked chicken breast, steak/chop bones with meat scraps left on, soft boiled eggs, cooked basmati rice and peanut butter or jam sandwiches. Only the best quality was given, however, the cost to provide so many possums with all of the aforementioned foods was astronomical and just when I thought I had the recipe right gastro intestinal tract (GIT) problems would arise. After attending a workshop conducted by Dr Anne Fowler in which she asked carers in attendance “why we were not feeding a more natural diet” I decided to take up the challenge; no longer would my ringtails, brushtails and gliders see anything remotely supplemental. -

List of Plants
Indigenous Plant Nursery Plant Species List The following plant list contains some of the local native plants that may be available from the Edendale Indigenous Plant Nursery. Availability can vary so please contact the nursery for specific and seasonal availability of plants. Contact details: [email protected] Phone (03) 9433 3703 30 Gastons Road, Eltham VIC 3091 Open 7 days per week, 9.30am to 4.30pm Trees Species Common Name Size (height x width) Acacia dealbata Silver Wattle 6 – 30m x 5 – 10m Acacia implexa Lightwood 5 – 15m x 4 – 7m Acacia pycnantha Golden Wattle 3 – 10m x 2 – 5m Acacia mearnsii Black Wattle 8 – 25m x 6 – 10m Acacia melanoxylon Blackwood 5 – 30m x 4 – 15m Allocasuarina littoralis Black Sheoke 4 – 8m x 2 – 5m Allocasuarina verticillata Drooping Sheoke 4 – 11m x 3 – 6m Banksia marginata Silver Banksia 1 – 10m x 1 – 5m Callitris rhomboidea Oyster Bay Pine 9 – 15 m high Eucalyptus blakelyi Blakely’s Red Gum 15 – 24m x 10 – 15m Eucalyptus camaldulensis River Red Gum 15 – 50m x 15 – 35m Eucalyptus goniocalyx Long-leaved Box 8 – 20m x 6 – 15m Eucalyptus leucoxylon Yellow Gum 10 – 20m x 6 – 20m Eucalyptus macrorhyncha Red Stringybark 10 – 35m x 10 – 20m Eucalyptus melliodora Yellow Box 10 – 30m x 8 – 25m Eucalyptus ovata Swamp Gum 8 – 30m x 8 – 20m Eucalyptus pauciflora Snow Gum 8 – 12m x 6 – 10m Eucalyptus polyanthemos Red Box 7 – 25m x 5 – 15m Eucalyptus radiata Narrow-leaved Peppermint 10 – 30m x 6 – 20m Eucalyptus rubida Candlebark Gum 10 – 25m x 10 – 20m Eucalyptus tricarpa Red Ironbark 10 – 30m x -

Indigenous Plant Guide
Local Indigenous Nurseries city of casey cardinia shire council city of casey cardinia shire council Bushwalk Native Nursery, Cranbourne South 9782 2986 Cardinia Environment Coalition Community Indigenous Nursery 5941 8446 Please contact Cardinia Shire Council on 1300 787 624 or the Chatfield and Curley, Narre Warren City of Casey on 9705 5200 for further information about indigenous (Appointment only) 0414 412 334 vegetation in these areas, or visit their websites at: Friends of Cranbourne Botanic Gardens www.cardinia.vic.gov.au (Grow to order) 9736 2309 Indigenous www.casey.vic.gov.au Kareelah Bush Nursery, Bittern 5983 0240 Kooweerup Trees and Shrubs 5997 1839 This publication is printed on Monza Recycled paper 115gsm with soy based inks. Maryknoll Indigenous Plant Nursery 5942 8427 Monza has a high 55% recycled fibre content, including 30% pre-consumer and Plant 25% post-consumer waste, 45% (fsc) certified pulp. Monza Recycled is sourced Southern Dandenongs Community Nursery, Belgrave 9754 6962 from sustainable plantation wood and is Elemental Chlorine Free (ecf). Upper Beaconsfield Indigenous Nursery 9707 2415 Guide Zoned Vegetation Maps City of Casey Cardinia Shire Council acknowledgements disclaimer Cardinia Shire Council and the City Although precautions have been of Casey acknowledge the invaluable taken to ensure the accuracy of the contributions of Warren Worboys, the information the publishers, authors Cardinia Environment Coalition, all and printers cannot accept responsi- of the community group members bility for any claim, loss, damage or from both councils, and Council liability arising out of the use of the staff from the City of Casey for their information published. technical knowledge and assistance in producing this guide. -

Wellington Shire Council Implementation of Coastal Strategy Land Capability Assessment
Wellington Shire Council Implementation of Coastal Strategy Land Capability Assessment March 2008 Contents 1. Background 1 1.1 Recent Strategy and Policy Decisions 1 1.2 Current Implementation Process 1 2. Methodology 2 2.1 Initial Methodology 2 2.2 Data Availability 2 2.3 Revised Methodology 3 3. Data 4 3.1 Data Sorting 4 3.2 Critical Data Sets 4 4. Analysis 12 4.1 Stage 1: Areas Inappropriate for Additional Dwellings 12 4.2 Stage 2: Capability Assessment for Dwelling Densities 12 4.3 Assigning Capability Scores for Dwelling Densities 13 5. Findings 18 5.1 Key Constraints to Development of Dwellings 18 5.2 Implications for Current Adopted Settlement Structure 21 5.3 Implications of Land Capability Assessment 23 5.4 Utilising the Outcomes of the Land Capability Assessment 23 Appendices A Flora and Fauna Report B Landscape Assessment Report C Table of Capability Scores for Dwelling Density Scenarios D Land Capability Mapping Implementation of Coastal Strategy i Land Capability Assessment 31/20287/138968 1. Background 1.1 Recent Strategy and Policy Decisions The Wellington Shire Council adopted the Wellington Coast Subdivision Strategy Option 4 – Nodal Urban settlement pattern at its meeting of 20th September, 2005 and the preparation of the Wellington Coast Subdivision Strategy: The Honeysuckles to Paradise Beach was finalised in February 2007. The Strategy was developed in accordance with the relevant coastal planning and management principles of the Victorian Coastal Strategy, 2002 prepared by the Victorian Coastal Council, and the Gippsland Coastal Board’s Coastal Action Plans. This strategy provides a preferred settlement structure for the Wellington Coast between Paradise Beach and The Honeysuckles, as a way to encourage development that responds appropriately to environmental values and community needs, and is consistent with established policy in the Victorian Coastal Strategy.