Plant Introduction МІЖНАРОДНИЙ НАУКОВИЙ ЖУРНАЛ • ЗАСНОВАНИЙ У 1999 Р
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

ZNIEFF Continentales : Liste Des Espèces De Flore Déterminantes En Région PACA
Actualisation de l’inventaire des Zones Naturelles d’Intérêt Écologique, Faunistique et Floristique (ZNIEFF) de Provence-Alpes-Côte d’Azur ZNIEFF continentales : liste des espèces de flore déterminantes en région PACA Version du 28/07/2016 Référentiel taxonomique : TAXREF v5.0 Les alismatales PHYLUM CLASSE ORDRE FAMILLE CD_REF RANG NOM_VALIDE Plantae Equisetopsida Alismatales Alismataceae 85486 ES Baldellia ranunculoides (L.) Parl., 1854 Plantae Equisetopsida Alismatales Alismataceae 160264 SSES Damasonium alisma Mill. subsp. polyspermum (Coss.) Maire Plantae Equisetopsida Alismatales Alismataceae 119860 ES Sagittaria sagittifolia L., 1753 Plantae Equisetopsida Alismatales Butomaceae 87136 ES Butomus umbellatus L., 1753 Plantae Equisetopsida Alismatales Hydrocharitaceae 103120 ES Hydrocharis morsus-ranae L., 1753 Plantae Equisetopsida Alismatales Hydrocharitaceae 128504 ES Vallisneria spiralis L., 1753 Plantae Equisetopsida Alismatales Juncaginaceae 141931 SSES Triglochin bulbosum subsp. barrelieri (Loisel.) Rouy, 1912 Plantae Equisetopsida Alismatales Juncaginaceae 127546 ES Triglochin maritimum L., 1753 Plantae Equisetopsida Alismatales Potamogetonaceae 81869 ES Althenia filiformis Petit, 1829 Plantae Equisetopsida Alismatales Potamogetonaceae 115228 ES Potamogeton alpinus Balb., 1804 Plantae Equisetopsida Alismatales Potamogetonaceae 115237 ES Potamogeton coloratus Hornem., 1813 Plantae Equisetopsida Alismatales Potamogetonaceae 115258 ES Potamogeton gramineus L., 1753 Plantae Equisetopsida Alismatales Potamogetonaceae 115296 ES Potamogeton -

Astereae, Asteraceae) Using Molecular Phylogeny of ITS
Turkish Journal of Botany Turk J Bot (2015) 39: 808-824 http://journals.tubitak.gov.tr/botany/ © TÜBİTAK Research Article doi:10.3906/bot-1410-12 Relationships and generic delimitation of Eurasian genera of the subtribe Asterinae (Astereae, Asteraceae) using molecular phylogeny of ITS 1, 2,3 4 Elena KOROLYUK *, Alexey MAKUNIN , Tatiana MATVEEVA 1 Central Siberian Botanical Garden, Siberian Branch of Russian Academy of Sciences, Novosibirsk, Russia 2 Institute of Molecular and Cell Biology, Siberian Branch of Russian Academy of Sciences, Novosibirsk, Russia 3 Theodosius Dobzhansky Center for Genome Bioinformatics, Saint Petersburg State University, Saint Petersburg, Russia 4 Department of Genetics & Biotechnology, Saint Petersburg State University, Saint Petersburg, Russia Received: 12.10.2014 Accepted/Published Online: 02.04.2015 Printed: 30.09.2015 Abstract: The subtribe Asterinae (Astereae, Asteraceae) includes highly variable, often polyploid species. Recent findings based on molecular methods led to revision of its volume. However, most of these studies lacked species from Eurasia, where a lot of previous taxonomic treatments of the subtribe exist. In this study we used molecular phylogenetics methods with internal transcribed spacer (ITS) as a marker to resolve evolutionary relations between representatives of the subtribe Asterinae from Siberia, Kazakhstan, and the European part of Russia. Our reconstruction revealed that a clade including all Asterinae species is paraphyletic. Inside this clade, there are species with unresolved basal positions, for example Erigeron flaccidus and its relatives. Moreover, several well-supported groups exist: group of the genera Galatella, Crinitaria, Linosyris, and Tripolium; group of species of North American origin; and three related groups of Eurasian species: typical Eurasian asters, Heteropappus group (genera Heteropappus, Kalimeris), and Asterothamnus group (genera Asterothamnus, Rhinactinidia). -

E. Biondi, L. Gubellini, M. Pinzi & S. Casavecchia the Vascular Flora Of
Fl. Medit. 22: 67-167 doi: 10.7320/FlMedit22.067 Version of Record published online on 28 December 2012 E. Biondi, L. Gubellini, M. Pinzi & S. Casavecchia The vascular flora of Conero Regional Nature Park (Marche, Central Italy) To Aldo J. B. Brilli-Cattarini, eminent botanist and expert resear- cher on the Marche region flora and par- ticularly on the Conero flora. He was the founder of the “Centro Ricerche Floristiche Marche” of Pesaro and Urbino Province. He has been a master for all of us even in the preparation of this work, with great gene rosity, he offered us his unique experience with enthusiasm. Abstract Biondi E., Gubellini L., Pinzi M. & Casavecchia, S.: The vascular flora of Conero Regional Nature Park (Marche, Central Italy). — Fl. Medit. 22: 67-167. 2012. — ISSN: 1120-4052 print- ed, 2240-4538 online. It is presented the vascular flora of Conero Regional Park. The paper starts with a brief presen- tation of the park territory (geography, geomorphology and bioclimate) that arises in the cen- tral Adriatic side of the Italian peninsula. The vegetation is briefly described thanks to the detailed phytosociological analysis carried out in the territory and to detailed maps on different scales. The description of vegetation allows indicating, for most of the species listed, their par- ticipation in the plant communities present in the area. The floristic list comprises 1169 entities, of specific and sub specific levels, that belong to 101 families and 507 genera. It also includes 64 species currently disappeared or not recently found in the study area, indicated by NP acronym (not present). -

Red List of Vascular Plants of the Czech Republic: 3Rd Edition
Preslia 84: 631–645, 2012 631 Red List of vascular plants of the Czech Republic: 3rd edition Červený seznam cévnatých rostlin České republiky: třetí vydání Dedicated to the centenary of the Czech Botanical Society (1912–2012) VítGrulich Department of Botany and Zoology, Masaryk University, Kotlářská 2, CZ-611 37 Brno, Czech Republic, e-mail: [email protected] Grulich V. (2012): Red List of vascular plants of the Czech Republic: 3rd edition. – Preslia 84: 631–645. The knowledge of the flora of the Czech Republic has substantially improved since the second ver- sion of the national Red List was published, mainly due to large-scale field recording during the last decade and the resulting large national databases. In this paper, an updated Red List is presented and compared with the previous editions of 1979 and 2000. The complete updated Red List consists of 1720 taxa (listed in Electronic Appendix 1), accounting for more then a half (59.2%) of the native flora of the Czech Republic. Of the Red-Listed taxa, 156 (9.1% of the total number on the list) are in the A categories, which include taxa that have vanished from the flora or are not known to occur at present, 471 (27.4%) are classified as critically threatened, 357 (20.8%) as threatened and 356 (20.7%) as endangered. From 1979 to 2000 to 2012, there has been an increase in the total number of taxa included in the Red List (from 1190 to 1627 to 1720) and in most categories, mainly for the following reasons: (i) The continuing human pressure on many natural and semi-natural habitats is reflected in the increased vulnerability or level of threat to many vascular plants; some vulnerable species therefore became endangered, those endangered critically threatened, while species until recently not classified may be included in the Red List as vulnerable or even endangered. -
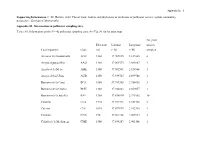
Supporting Information. C. M. Herrera. 2020. Flower Traits, Habitat, and Phylogeny As Predictors of Pollinator Service: a Plant Community Perspective
Appendix S1 – 1 Supporting Information. C. M. Herrera. 2020. Flower traits, habitat, and phylogeny as predictors of pollinator service: a plant community perspective. Ecological Monographs. Appendix S1. Information on pollinator sampling sites. TABLE S1. Information on the N = 42 pollinator sampling sites. See Fig. S1 for location map. No. plant Elevation Latitude Longitude species Local toponym Code (m) (º N) (º W) sampled Arenales del Guadalentín AGU 1360 37.909835 2.837405 4 Arroyo Aguaderillos AAG 1180 37.961573 2.883887 1 Arroyo de la Mesa AME 1100 37.902591 2.928966 1 Arroyo de los Ubios AUB 1250 37.939725 2.899766 1 Barranco de la Canal BCA 1580 37.789385 2.956438 3 Barranco de la Charca BCH 1360 37.942843 2.859057 1 Barranco de la Juanfría BJU 1380 37.836814 2.975302 10 Calarilla CLA 1710 37.944316 2.851708 2 Calerón CLE 1075 37.897973 2.942710 1 Cantalar CAN 770 37.964158 2.907974 3 Cañada de la Medianega CME 1580 37.894287 2.903146 1 Appendix S1 – 2 Cañada Pajarera CPA 1640 37.927456 2.805615 1 Collado del Calvario CCA 1420 37.950202 2.885484 1 Collado Zamora CZA 1420 37.839329 2.999615 2 Coto del Valle CVA 850 37.930542 2.928517 7 Cuesta del Bazar CBA 1330 37.906523 2.907470 1 Cuevas Bermejas CBE 1160 37.966959 2.851117 5 Fuente Bermejo FBE 1550 37.932268 2.840672 14 Fuente la Reina FRE 1460 37.941698 2.833632 2 Guadalentín GUA 1310 37.900404 2.836944 8 Hoyos de Muñoz HMU 1110 37.984831 2.876597 3 La Cabrilla LCA 1600 37.932364 2.780637 5 La Fresnedilla LFR 1080 37.980387 2.855875 2 La Mesa LME 1610 37.889823 2.913788 2 Las -

Modifications Taxonomiques Des Espèces
Modifications taxonomiques des espèces Migration de TAXREF version 3.0 (20/09/2010) à la version 4.0 (12/10/2011) (source: http://inpn.mnhn.fr/isb/programmes/fr/taxrefPres.jsp) TAXREF v .3.0 TAXREF v .4.0 R: Animalia Animalia E: Arthropoda Arthropoda C: Arachnida Arachnida O: Araneae Araneae F: Sparassidae Sparassidae - Micrommata virescens ornatum - Micrommata virescens ornata C: Chilopoda Chilopoda O: Geophilomorpha Geophilomorpha F: Geophilidae Geophilidae - Geophilus linearis - Stenotaenia linearis C: Insecta Insecta O: Coleoptera Coleoptera F: Dytiscidae Dytiscidae - Eretes sticticus - Eretes griseus O: Diptera Diptera F: Chironomidae Chironomidae - Metriocnemus hirticollis - Metriocnemus eurynotus F: Ephydridae Ephydridae - Teichomyza fusca - Scatella (Teichomyza) fusca F: Syrphidae Syrphidae - Eristalinus aeneus - Eristalis aeneus O: Hymenoptera Hymenoptera F: Ichneumonidae Ichneumonidae - Diplazon multicolor - Diplazon annulatus O: Lepidoptera Lepidoptera F: Hesperiidae Hesperiidae - Carcharodus boeticus - Carcharodus baeticus - Carcharodus floccifera - Carcharodus floccifer F: Lycaenidae Lycaenidae - Cupido carswelli - Cupido minimus - Glaucopsyche alcon - Maculinea alcon alcon - Glaucopsyche arion - Maculinea arion - Glaucopsyche iolas - Iolana iolas - Glaucopsyche nausithous - Maculinea nausithous - Glaucopsyche rebeli - Maculinea alcon rebeli - Glaucopsyche teleius - Maculinea teleius - Neozephyrus quercus - Quercusia quercus - Plebeius agestis - Aricia agestis - Plebeius argus - Plebejus argus - Plebeius argyrognomon - Plebejus -

Svensk Botanisk Tidskrift Otaniska Otaniska Öreningen Venska Venska Sveriges Kalkmossor Sveriges S B F Årets Mossa: Briljantmossa Mossa: Årets
Styv kalkmossa och alvarkalkmossa Svensk Många botanister känner På bilden ser vi styv kalkmossa har hittats i Sverige helt nyli säkert till kruskalkmossa till vänster, gen, och det kan finnas fler. Läs Tortella rigens Svenska som en bra kalk tillsammans med den betyd mer om våra kalkmossor på T. tortuosa Botaniska indikator, men hur många vet ligt mindre alvarkalkmossan sid. 94. Botanisk Föreningen att vi har minst ytterligare sju T. densa. FOTO: Lars Hedenäs. – Gotland, arter i släktet kalkmossor i Alla arter i släktet växer i Hellvi s:n, mellan Sudergårde landet? spännande miljöer, några arter och Lörge, 13 maj 2013. Tidskrift Svensk Botanisk Tidskrift Volym 109: Häfte 2, 2015 109(2): 65–128 (2015) 109(2): Sveriges kalkmossor Årets mossa: Briljantmossa Nya arter och namn i Nordens flora innehåll våra regionala föreningar Hallands Botaniska Förening Hallands Botaniska Förening har ungefär 300 Varje år ger vi ut en betalande medlemmar, Vårt mål är att öka innehållsrik botanisk kunskaperna om den botaniska mångfalden utflyktsguide för Halland i Halland, att arbeta för bevarandet av denna i samband med det nya mångfald och att stimulera intresset för bota programmet för årets nik i allmänhet. aktiviteter. Wiveka Johansson Niklas Lönnell : : Föreningen bildades 2002 på initiativ av FOTO FOTO bland andra Kjell Georgson som var huvud Där nere någonstans Sälgens blomning och ansvarig för Hallands flora som kom 1997. gömmer sig den mycket honungsbinas första drag. Vi ordnar varje år ett stort antal exkursioner 120lilla millimeter mossan. 112 Två klassiska vårtecken. över hela landskapet med fokus på kärlväx Men Niklas Lönnell hittade den ändå, i Var med och rapportera vårtecken du ter, mossor och svampar. -

Composition and Antimicrobial Activity of the Essential Oil from Galatella Linosyris (L.) Rchb
J. Serb. Chem. Soc. 77 (5) 619–626 (2012) UDC *Galatella linosyris:665.52/.54:615.28–188 JSCS–4295 Original scientific paper Composition and antimicrobial activity of the essential oil from Galatella linosyris (L.) Rchb. f. (Asteraceae) DEJAN GOĐEVAC1*, LJUBODRAG VUJISIĆ2, IVAN VUČKOVIĆ2, VLATKA VAJS1, MARINA SOKOVIĆ3, PETAR D. MARIN4# and VELE TEŠEVIĆ2 1Institute of Chemistry, Technology and Metallurgy, Njegoševa 12, University of Belgrade, 11000 Belgrade, Serbia, 2Faculty of Chemistry, University of Belgrade, Studentski trg 16, P. O. Box 158, 11000 Belgrade, Serbia, 3Institute for Biological Research “Siniša Stanković”, University of Belgrade, Bulevar despota Stefana 142, 11000 Belgrade, Serbia and 4Faculty of Biology, University of Belgrade, Institute of Botany and Botanical Garden “Jevremovac”, Studentski trg 16, 11000 Belgrade, Serbia (Received 15 September, revised 15 December 2011) Abstract: An investigation of the chemical composition and antimicrobial ac- tivity of the essential oil of Galatella linosyris is presented. The chemical anal- ysis (GC/MS, NMR) showed that sabinene (40 %), β-pinene (35.5 %), α-pi- nene (4.5 %), limonene (4 %), γ-muurolene (4 %), and (E)-caryophyllene (3.3 %) were dominant components in this oil. Microdilution assays were used to evaluate the minimum inhibitory concentration (MIC) and the minimum bac- tericidal/fungicidal concentrations (MBC/MFC). G. linosyris essential oil exhi- bited better antibacterial activity against some of the tested bacteria than anti- fungal activity. Keywords: Galatella linosyris; -

First Updated Checklist of the Vascular Flora of Andalusia (S of Spain), One of the Main Biodiversity Centres in the Mediterranean Basin
Phytotaxa 339 (1): 001–095 ISSN 1179-3155 (print edition) http://www.mapress.com/j/pt/ PHYTOTAXA Copyright © 2018 Magnolia Press Monograph ISSN 1179-3163 (online edition) https://doi.org/10.11646/phytotaxa.339.1.1 PHYTOTAXA 339 First updated checklist of the vascular flora of Andalusia (S of Spain), one of the main biodiversity centres in the Mediterranean Basin MIGUEL CUETO1,*, MANUEL MELENDO2, ESTHER GIMÉNEZ1, JULIÁN FUENTES3, ENRIQUE LÓPEZ CARRIQUE4 & GABRIEL BLANCA5 1 Departamento de Biología y Geología, CECOUAL, Universidad de Almería, Crta. Sacramento s/n, La Cañada de S. Urbano, ES- 04120 Almería, Spain; e-mail: [email protected], [email protected] 2 Departamento de Biología Animal, Biología Vegetal y Ecología. Facultad de Ciencias Experimentales, Universidad de Jaén, Campus Las Lagunillas s/n, ES-23071 Jaén, Spain; e-mail: [email protected] 3 C/ Castillo 5, bajo F, ES-18140 La Zubia, Granada, Spain; e-mail: [email protected] 4 Departamento de Educación, CECOUAL, Universidad de Almería, Crta. Sacramento s/n, La Cañada de S. Urbano, ES-04120 Almería, Spain; e-mail: [email protected] 5 Departamento de Botánica, Facultad de Ciencias, Universidad de Granada, C/ Fuentenueva s/n, ES-18001 Granada, Spain; e-mail: [email protected] *author for correspondence Magnolia Press Auckland, New Zealand Accepted by Manuel B. Crespo: 24 Dec. 2017; published: 20 Feb. 2018 MIGUEL CUETO, MANUEL MELENDO, ESTHER GIMÉNEZ, JULIÁN FUENTES, ENRIQUE LÓPEZ CARRIQUE & GABRIEL BLANCA First updated checklist of the vascular flora of Andalusia (S of Spain), one of the main biodiversity centres in the Mediterranean Basin (Phytotaxa 339) 95 pp.; 30 cm. -

Detail Galatella Linosyris 17. 7. 2021
Dump from Pladias.cz portal 21.9.2021 Galatella linosyris Distribution Habitus and growth type Height [m]: 0.15–0.4 Growth form: clonal herb Life form: hemicryptophyte Life strategy: CSR – competitor/stress-tolerator/ruderal Life strategy (Pierce method based on leaf traits): S/SR Life strategy (Pierce method, C-score): 6.9 % Life strategy (Pierce method, S-score): 78.2 % Life strategy (Pierce method, R-score): 15 % Leaf Leaf presence and metamorphosis: leaves present, not modified Leaf arrangement (phyllotaxis): alternate Leaf shape: simple – entire Stipules: absent Petiole: absent Leaf life span: summer green Leaf anatomy: scleromorphic, mesomorphic Flower Flowering period [month]: July-September © 2014–2021 Pladias – Citation: Pladias – Database of the Czech Flora and Vegetation. www.pladias.cz Dump from Pladias.cz portal 21.9.2021 Flowering phase: 8 Clematis vitalba-Galium sylvaticum (mid-summer) Flower colour: yellow Flower symmetry: actinomorphic Perianth type: calyx reduced, corolla present Perianth fusion: fused Shape of the sympetalous corolla or syntepalous perianth: tubular Calyx fusion: pappus Inflorescence type: corymbothyrsus ex anthodiis compositus Dicliny: synoecious Generative reproduction type: allogamy self-incompatibility, facultative allogamy Pollination syndrome: insect-pollination, geitonogamy Fruit, seed and dispersal Fruit type: dry fruit – achene/cypsela/samara Fruit colour: brown Reproduction type: by seed/spores and vegetatively Dispersal unit (diaspore): fruit, infrutescence or its part Dispersal strategy: -

Bulletin De La Société Botanique Du Centre-Ouest - Nouvelle Série - Tome 51 - 2020 2020 - Bull
Numéros spéciaux publiés par la SBCO • Clés de détermination des Bryophytes de la région Poitou-Charentes- Vendée, par R.B. PIERROT (1974 – n° 1) • Matériaux pour une étude floristique et phytosociologique du Limousin occidental : Forêt de Rochechouart et secteurs limitrophes (Haute-Vienne), par H. BOUBY. 134 p. (1978 – n° 2) • Les Discomycètes de France d’après la classification de Boudier, par L.-J. GRELET, 1979. 709 pages (réédition). (1979 – n° 3) • La vie dans les dunes du Centre-Ouest : flore et faune. Ouvrage collectif. 213 p. (1980 – n° 4) • Les Bryophytes du Centre-Ouest, par R.B. PIERROT. 120 p. (réimpr. 2005 – n° 5) • Contribution à l’étude botanique de la haute et moyenne vallée de la Vienne (phytogéographie et phytosociologie), par M. BOTINEAU. VI + 352 p. En annexe 40 tableaux phytosociologiques. (1982 – n° 6) BULLETIN • Likenoj de Okcidenta Europo. Ilustrita determinlibro (Lichens d’Europe occidentale. Flore illustrée. Rédigée en espéranto), par G. CLAUZADE et C. ROUX. 893 p. (1985 – n° 7) • Index synonymique de la flore des régions occidentales de la France (Plantes vasculaires), par le Professeur P. DUPONT. 246 p. (1986 – n° 8) • La végétation de la Basse-Auvergne, par F. BILLY. 416 p. (1988 – n° 9) de la • Les Festuca de la flore de France (Corse comprise), par M. KERGUÉLEN et F. PLONKA. Avant-propos du Prof. J. LAMBINON. 368 p. (1989- n° 10) • Phytosociologie et écologie des forêts de Haute-Normandie. Leur place dans le contexte sylvatique ouest-européen, par J. BARDAT. 376 p + 85 tableaux phytosociologiques. (1993 – n° 11) SOCIÉTÉ BOTANIQUE • Pelouses et ourlets du Berry, par R. -

TUL Herbarium: Collections of Vascular Plants of Tula Oblast, Russia
Biodiversity Data Journal 8: e61454 doi: 10.3897/BDJ.8.e61454 Data Paper TUL Herbarium: collections of vascular plants of Tula Oblast, Russia Tatyana Yu. Svetasheva‡, Alexey P. Seregin§ ‡ Tula State Lev Tolstoy Pedagogical University, Tula, Russia § Lomonosov Moscow State University, Moscow, Russia Corresponding author: Tatyana Yu. Svetasheva ([email protected]) Academic editor: Ivan Chadin Received: 28 Nov 2020 | Accepted: 21 Dec 2020 | Published: 24 Dec 2020 Citation: Svetasheva TYu, Seregin AP (2020) TUL Herbarium: collections of vascular plants of Tula Oblast, Russia. Biodiversity Data Journal 8: e61454. https://doi.org/10.3897/BDJ.8.e61454 Abstract Background TUL Herbarium presents collections from Tula Oblast stored at the Tula State Lev Tolstoy Pedagogical University, Russia, which is an educational and scientific institution that supports various types of scientific activities, including research on biodiversity and nature conservation. The university is a holder of some biological collections, such as herbarium of vascular plants, mosses and fungi collected mainly throughout Tula Oblast and from adjacent regions. New information The collections of vascular plants (9,000 specimens) were imaged in December 2019 and January 2020. Databasing and georeferencing of the specimens from the TUL Herbarium was performed by the staff members of the Tula State Lev Tolstoy Pedagogical University and Tula Local History Museum. Digital collections of the TUL Herbarium are fully available in the Moscow Digital Herbarium (https://plant.depo.msu.ru/) and GBIF (https://doi.org/ 10.15468/ca08cm). © Svetasheva T, Seregin A. This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.