American Beech in Coastal New England: Forest History and Dynamics
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Addressing Effects of Offshore Wind Development on Bat Species in the Northeastern United States
University of Massachusetts Amherst ScholarWorks@UMass Amherst Doctoral Dissertations Dissertations and Theses November 2018 NOT GONE WITH THE WIND: Addressing Effects of Offshore Wind Development on Bat Species in the Northeastern United States Zara Rae Dowling University of Massachusetts Amherst Follow this and additional works at: https://scholarworks.umass.edu/dissertations_2 Part of the Other Ecology and Evolutionary Biology Commons, Terrestrial and Aquatic Ecology Commons, and the Zoology Commons Recommended Citation Dowling, Zara Rae, "NOT GONE WITH THE WIND: Addressing Effects of Offshore Wind Development on Bat Species in the Northeastern United States" (2018). Doctoral Dissertations. 1427. https://doi.org/10.7275/12644119 https://scholarworks.umass.edu/dissertations_2/1427 This Open Access Dissertation is brought to you for free and open access by the Dissertations and Theses at ScholarWorks@UMass Amherst. It has been accepted for inclusion in Doctoral Dissertations by an authorized administrator of ScholarWorks@UMass Amherst. For more information, please contact [email protected]. NOT GONE WITH THE WIND: ADDRESSING EFFECTS OF OFFSHORE WIND DEVELOPMENT ON BAT SPECIES IN THE NORTHEASTERN UNITED STATES A Dissertation Presented by ZARA RAE DOWLING Submitted to the Graduate School of the University of Massachusetts Amherst in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY September 2018 Environmental Conservation © Copyright by Zara Rae Dowling 2018 All Rights Reserved NOT GONE WITH THE -

Cuttyhunk-Nantucket 24-Quadrangle Area of Cape Cod and Islands, Southeast Massachusetts
Prepared in cooperation with the Commonwealth of Massachusetts Office of the State Geologist and Executive Office of Energy and Environmental Affairs Surficial Geologic Map of the Pocasset-Provincetown- Cuttyhunk-Nantucket 24-Quadrangle Area of Cape Cod and Islands, Southeast Massachusetts Compiled by Byron D. Stone and Mary L. DiGiacomo-Cohen Open-File Report 2006-1260-E U.S. Department of the Interior U.S. Geological Survey U.S. Department of the Interior KEN SALAZAR, Secretary U.S. Geological Survey Marcia K. McNutt, Director U.S. Geological Survey, Reston, Virginia: 2009 For product and ordering information: World Wide Web: http://www.usgs.gov/pubprod Telephone: 1-888-ASK-USGS For more information on the USGS—the Federal source for science about the Earth, its natural and living resources, natural hazards, and the environment: World Wide Web: http://www.usgs.gov Telephone: 1-888-ASK-USGS Suggested citation: Stone, B.D., and DiGiacomo-Cohen, M.L., comps., 2009, Surficial geologic map of the Pocasset Provincetown-Cuttyhunk-Nantucket 24-quadrangle area of Cape Cod and Islands, southeast Massachusetts: U.S. Geological Survey Open-File Report 2006-1260-E. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government. Although this report is in the public domain, permission must be secured from the individual copyright owners to reproduce any copyrighted materials contained within this report. Cover figure. Photograph of eroding cliffs at Gay Head on Martha’s Vineyard (source: -

Dukes County, Massachusetts (All Jurisdictions)
DUKES COUNTY, MASSACHUSETTS (ALL JURISDICTIONS) COMMUNITY NAME COMMUNITY NUMBER AQUINNAH, TOWN OF 250070 CHILMARK, TOWN OF 250068 EDGARTOWN, TOWN OF 250069 GOSNOLD, TOWN OF 250071 OAK BLUFFS, TOWN OF 250072 TISBURY, TOWN OF 250073 WEST TISBURY, TOWN OF 250074 Dukes County Federal Emergency Management Agency FLOOD INSURANCE STUDY NUMBER 25007CV000B NOTICE TO FLOOD INSURANCE STUDY USERS Communities participating in the National Flood Insurance Program have established repositories of flood hazard data for floodplain management and flood insurance purposes. This Flood Insurance Study (FIS) may not contain all data available within the repository. It is advisable to contact the community repository for any additional data. Selected Flood Insurance Rate Map panels for the community contain information that was previously shown separately on the corresponding Flood Boundary and Floodway Map panels (e.g., floodways, cross sections). In addition, former flood hazard zone designations have been changed as follows: Old Zone New Zone A1 through A30 AE V1 through V30 VE B X C X Part or all of this Flood Insurance Study may be revised and republished at any time. In addition, part of this Flood Insurance Study may be revised by the Letter of Map Revision process, which does not involve republication or redistribution of the Flood Insurance Study. It is, therefore, the responsibility of the user to consult with community officials and to check the community repository to obtain the most current Flood Insurance Study components. Initial Countywide FIS -

Favorite Cruising Spots in Buzzard's Bay and Vineyard Sound Cuttyhunk Island—Typically a 1.5 Hour Sail on a Close Reach; A
Favorite Cruising spots in Buzzard’s Bay and Vineyard Sound Cuttyhunk Island—typically a 1.5 hour sail on a close reach; a tiny island and crowded anchorage but a fun and very popular place to stop and walk around, stop for lunch or stay overnight. Inner Harbor is well-protected with both moorings and small anchorage available. Moorings also available along beach outside of breakwater between harbor and Nashawena, and good holding ground for anchoring outside of breakwater if anchorage is full. Lovely view of Elizabeth Island chain from top of hill. There is a market on the way up the hill from the main dock on your left. Also a store with Cuttyhunk “stuff” on the right after the dock. Also a tiny shack to the left which has AMAZING clothing and jewelry—bring cash! Fun local pizza called SOPRANO’s (“pizza to die for”). The best kept secret is “Cuttyhunk Fishing Club” which serves THE BEST breakfast al fresco on their porch on a hill overlooking the Elizabeth Islands and Martha’s Vineyard. Quick’s Hole—The safest passage between Buzzard’s Bay and Vineyard Sound. Quicks is nestled between Pasque and Nashawena Islands (two more Elizabeth Islands) and as such is normally protected from wind and seas. There’s a lovely white sand beach with dunes and lots of good anchorage fairly close up. Great lunch spot. The beach can be closed at random by the owners of Naushon Island however. Lambert’s Cove on North side of Martha’s Vineyard on Vineyard Sound side. Nice beach with dunes for a swim and picnic lunch in normal SW wind. -

FY2005 Annual Report
AnnAnnualual ReportReport 20052005 MassachusettsMassachusetts DivisionDivision ofof FisheriesFisheries && WildlifeWildlife Annual Report 2005 Massachusetts Division of fisheries & WilDlife Wayne F. MacCallum Director Jack Buckley Rob Deblinger, Ph.D. Deputy Director Deputy Director Administration Field Operations Table of Contents The Board Reports.............................................................................................4 Fisheries.............................................................................................................8 Wildlife.............................................................................................................17 Landowner Incentive Program .......................................................................29 Natural Heritage & Endangered Species Program.........................................31 Information & Education................................................................................37 Hunter Education............................................................................................40 District Reports ...............................................................................................45 Wildlife Lands ..................................................................................................55 Federal Aid Program Administration..............................................................61 Maintenance and Development.......................................................................63 Legislative Report............................................................................................64 -

USGS 7.5-Minute Image Map for Naushon Island, Massachusetts
U.S. DEPARTMENT OF THE INTERIOR NAUSHON ISLAND QUADRANGLE U. S. GEOLOGICAL SURVEY MASSACHUSETTS-DUKES CO. O C 7.5-MINUTE SERIES L O O T C IS R 7ES0°52'30" 50' 47'30" 70°45' B K U 3 000m 3 3 3 3 3 3 3 3 D 44 E 45 46 48 49 50 51 52 53 860 000 FEET 41°30' 41°30' K N U H Y T T U G H IN C S - S O D R R C FO D Y E R B R FE W E N 4595 4595000mN Mount 2 64C0 a00r0y Silver Beach FEET 100 0 0 1 45 0 94 0 1 5 0 4594 50 South Bluff " Elizabeth Islands " Kettle " " " " " Cove " " " " " Rams Head Tarpaulin 4593 H î " î H " " " " " ove " C H " "" " " î î """ î "H " " H " 4593 50 50 Blaney 0 nd 5 Po Naushon Island " " î H " H " H " H " " î " î î " H H " " Hî î î î î " î " Hî H î î H " î H H î " H " î " " î H " H î î " " H " H î " H ove Light î C î in î î ul î H Tarpa H H " H " î H î H H î " H î î H î " " H " H " " H H H "" î H î " " î H î ch H a î e î î B H nt " H H ce î es î r î C H H H " H î î " î " " H " î H " H H 5 " î H " î " î î H î î î H H î H H H H î H H " 0 H î " î î " " " î " " " " î H " H " " î î H î " î H î î " î " H H H " " H î H î " î î " î î H H î î î " H î H H î " H H H î " " î î î î H " H î H H " H " H î î " " î " î H î H " " î î " H î " " H î H î " " î î " " î " H H " î " î " " H " " H î " " " " H " î H H î " î " H H H î H " î H " î " î H H H î î î î " î H 10 î 0 î H " î H H " H î " " î " H î î " " H î H H " " " " " î 45 î " " " î H î H " î î 92 " H î H H H" H " " " H H H H î î " " î H î " î " î " î " H î î H î î î î H î H " " " î " " H " î î H H î " H " î H" H" H î " î H " " H" " " " " î " î î " î H î "" " " î H î " î H " H î " " î -

Annual Report 2018
Massachusetts Division of Fisheries & Wildlife 2018 Annual Report 147 Annual Report 2018 Massachusetts Division of Fisheries & Wildlife Jack Buckley Director (July 2017–May 2018) Mark S. Tisa, Ph.D., M.B.A. Acting Director (May–June 2018) 149 Table of Contents 2 The Board Reports 6 Fisheries 42 Wildlife 66 Natural Heritage & Endangered Species Program 82 Information & Education 95 Archivist 96 Hunter Education 98 District Reports 124 Wildlife Lands 134 Federal Aid 136 Staff and Agency Recognition 137 Personnel Report 140 Financial Report Appendix A Appendix B About the Cover: MassWildlife staff prepare to stock trout at Lake Quinsigamond in Worcester with the help of the public. Photo by Troy Gipps/MassWildlife Back Cover: A cow moose stands in a Massachusetts bog. Photo by Bill Byrne/MassWildlife Printed on Recycled Paper. ELECTRONIC VERSION 1 The Board Reports Joseph S. Larson, Ph.D. Chairperson Overview fective April 30, 2018, and the Board voted the appoint- ment of Deputy Director Mark Tisa as Acting Director, The Massachusetts Fisheries and Wildlife Board con- effective Mr. Buckley’s retirement. The Board -mem sists of seven persons appointed by the Governor to bers expressed their gratitude and admiration to the 5-year terms. By law, the individuals appointed to the outgoing Director for his close involvement in develop- Board are volunteers, receiving no remuneration for ing his staff and his many accomplishments during his their service to the Commonwealth. Five of the sev- tenure, not only as Director but over his many years as en are selected on a regional basis, with one member, Deputy Director in charge of Administration, primarily by statute, representing agricultural interests. -

Recommended Time of Year Restrictions (Toys) for Coastal Alteration Projects to Protect Marine Fisheries Resources in Massachusetts
Massachusetts Division of Marine Fisheries Technical Report TR-47 Recommended Time of Year Restrictions (TOYs) for Coastal Alteration Projects to Protect Marine Fisheries Resources in Massachusetts N. T. Evans, K. H. Ford, B. C. Chase, and J. J. Sheppard Commonwealth of Massachusetts Executive Office of Energy and Environmental Affairs Department of Fish and Game Massachusetts Division of Marine Fisheries Technical Report Technical April 2011 Revised January 2015 Massachusetts Division of Marine Fisheries Technical Report Series Managing Editor: Michael P. Armstrong Scientific Editor: Bruce T. Estrella The Massachusetts Division of Marine Fisheries Technical Reports present information and data pertinent to the management, biology and commercial and recreational fisheries of anadromous, estuarine, and marine organisms of the Commonwealth of Massachusetts and adjacent waters. The series presents information in a timely fashion that is of limited scope or is useful to a smaller, specific audience and therefore may not be appropriate for national or international journals. Included in this series are data summaries, reports of monitoring programs, and results of studies that are directed at specific management problems. All Reports in the series are available for download in PDF format at: http://www.mass.gov/marinefisheries/publications/technical.htm or hard copies may be obtained from the Annisquam River Marine Fisheries Station, 30 Emerson Ave., Gloucester, MA 01930 USA (978-282-0308). TR-1 McKiernan, D.J., and D.E. Pierce. 1995. The Loligo squid fishery in Nantucket and Vineyard Sound. TR-2 McBride, H.M., and T.B. Hoopes. 2001. 1999 Lobster fishery statistics. TR-3 McKiernan, D.J., R. Johnston, and W. -

Massachusetts Summary of Proposed Changes
U.S. Fish & Wildlife Service John H. Chafee Coastal Barrier Resources System (CBRS) Unit C00, Clark Pond, Massachusetts Summary of Proposed Changes Type of Unit: System Unit County: Essex Congressional District: 6 Existing Map: The existing CBRS map depicting this unit is: ■ 025 dated October 24, 1990 Proposed Boundary Notice of Availability: The U.S. Fish & Wildlife Service (Service) opened a public comment period on the proposed changes to Unit C00 via Federal Register notice. The Federal Register notice and the proposed boundary (accessible through the CBRS Projects Mapper) are available on the Service’s website at www.fws.gov/cbra. Establishment of Unit: The Coastal Barrier Resources Act (Pub. L. 97-348), enacted on October 18, 1982 (47 FR 52388), originally established Unit C00. Historical Changes: The CBRS map for this unit has been modified by the following legislative and/or administrative actions: ■ Coastal Barrier Improvement Act (Pub. L. 101-591) enacted on November 16, 1990 (56 FR 26304) For additional information on historical legislative and administrative actions that have affected the CBRS, see: https://www.fws.gov/cbra/Historical-Changes-to-CBRA.html. Proposed Changes: The proposed changes to Unit C00 are described below. Proposed Removals: ■ One structure and undeveloped fastland near Rantoul Pond along Fox Creek Road ■ Four structures and undeveloped fastland located to the north of Argilla Road and east of Fox Creek Proposed Additions: ■ Undeveloped fastland and associated aquatic habitat along Treadwell Island Creek, -
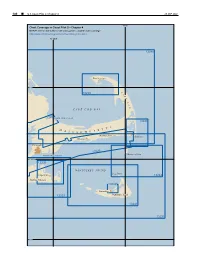
Outer Cape Cod and Nantucket Sound
186 ¢ U.S. Coast Pilot 2, Chapter 4 26 SEP 2021 70°W Chart Coverage in Coast Pilot 2—Chapter 4 NOAA’s Online Interactive Chart Catalog has complete chart coverage http://www.charts.noaa.gov/InteractiveCatalog/nrnc.shtml 70°30'W 13246 Provincetown 42°N C 13249 A P E C O D CAPE COD BAY 13229 CAPE COD CANAL 13248 T S M E T A S S A C H U S Harwich Port Chatham Hyannis Falmouth 13229 Monomoy Point VINEYARD SOUND 41°30'N 13238 NANTUCKET SOUND Great Point Edgartown 13244 Martha’s Vineyard 13242 Nantucket 13233 Nantucket Island 13241 13237 41°N 26 SEP 2021 U.S. Coast Pilot 2, Chapter 4 ¢ 187 Outer Cape Cod and Nantucket Sound (1) This chapter describes the outer shore of Cape Cod rapidly, the strength of flood or ebb occurring about 2 and Nantucket Sound including Nantucket Island and the hours later off Nauset Beach Light than off Chatham southern and eastern shores of Martha’s Vineyard. Also Light. described are Nantucket Harbor, Edgartown Harbor and (11) the other numerous fishing and yachting centers along the North Atlantic right whales southern shore of Cape Cod bordering Nantucket Sound. (12) Federally designated critical habitat for the (2) endangered North Atlantic right whale lies within Cape COLREGS Demarcation Lines Cod Bay (See 50 CFR 226.101 and 226.203, chapter 2, (3) The lines established for this part of the coast are for habitat boundary). It is illegal to approach closer than described in 33 CFR 80.135 and 80.145, chapter 2. -

Finding Aid to Family Collections
FALMOUTH HISTORICAL SOCIETY GUIDE TO COLLECTIONS OF FAMILY PAPERS Bodfish Family Collection 1803-1907 The bulk of this collection derives from the activities of Captain William Bodfish (1789- 1824) and his son Robinson Crocker Bodfish (1818-1912). The papers of Captain Bodfish, a trader, who shipped goods from Puerto Rico, St. Pierre, Martinique, and St. John, consists of correspondence with other shipping merchants (ca.1815-1824); receipts, invoices, and freight lists for goods shipped on brigs Fame, Caroline, Franklin, and Clarion, and schooner Leon (ca.1819-1824). In addition there are papers (1824-1828) relating to William’s estate for which his wife, Mary Crocker Bodfish, was executrix. The papers of Robinson Crocker Bodfish reflect his interest as an amateur local historian and his ambitions for the development in the Ocean Vistas and Falmouth Heights areas. They consist of correspondence (ca.1842-1905) relating to real estate developments, maps and plans for the development of the village of Oak Bluffs in Martha’s Vineyard; letters (ca.1859-1872) from Frederick Freeman, author of The History of Cape Cod (1858); a scrapbook of clippings of Falmouth history (1900-1904); and a compilation of data on the whaling industry of Falmouth. Completing the collection are papers and/or photographs of Abigail Fessenden (Mrs. William) Bodfish (1764-1852); Deborah Frances Bodfish Woodberry (1820-1851); H.H. Bodfish; the Reverend J.P. Bodfish; Joseph C. Bodfish; Lydia Soule (Mrs. Robinson Crocker) Bodfish; Mary Crocker (Mrs. William) Bodfish (1788-1847); Sumner H. Bodfish; and another William Bodfish (1758-1809). 2 boxes. See also Genealogy Collection. -

Executive. *1 General Post Office
EXECUTIVE. *1 GENERAL POS? OFFICE. P. Mast. Qen. Clks. kc. GENERAL POST OFFICE. Persons employed in the General Post Office, with the annual compensation ofeach> from the 1st day ofOcidber, 1829. NAMES AND OFFICES: POSTMASTER GENERAL. William T.Barry,.. ASSISTANT POSTMASTERS' GENERAL. Charles, K. Gardner,. S.elah R. Hobbie, .. CHIEF CLERK. Obadiab. B..Brown,........... CLERKS. Thomas B. Dyer,.:......, Joseph W. Haiid,. ; John Suter,.'.;; "... John McLeod, '....'. William G. Elliot, Michael T. Simpson,... Nicholas Tastet, David Saunders,... Rchard; Dement, Willing Blair, Thomas Arbuckle, Josiah f. Caldwell, "Joseph Haskell...... Samuel' Fitzhugh, William C.Ellison,.."... William Deming, Hyilliaift Cl'Lipscomb,. 'Thomas B; Addison,.:.'.' Matthias Ross, Davidj^oones, JfctitUy, Sinlpson,.....'.. A EXECUTIVE. GENERAL POST OFFICE. P Mast. Gen. Clks.kc. Compen NAMES AND OFFICES. sation &c. D. C. Grafton D. Hanson, 1000 00 Walter D. Addison,.. 1000 00 Andrew McD. Jackson,.... 1000 00 Arthur Nelson, 1000 00 John W. Overton, 1000 00 Henry S. Handy, Samuel Gwin, 1000 0® LemueLW. Ruggles, 1000 00 George S. Douglass, 1000 CO Preston S. Loughborough,. 1000 00 Francis G. Blackford, 1000 00 John G. Whitwell, 800 00 Thomas E. Waggoman,.... 800 0» John A Collins, Joseph Sherrill, 800 00 John F. Boone, 800 00 John G. Johnson, 800 0t John L. Storer, 800 0« William French, 800 09 James H. Doughty, 800 00 James Coolidge,., 800 00 Charles S. Williams, EdmundF. Brown, 800 00 Alexander H. Fitzhugh,.... 800 00 800 00 FOR OPENING DEAD LETTERS. 800 00 500 00 Charles Bell, 400 00 William Harvey,. 400 00 MESSENGER. Joseph Borrows, 700 0» ASSISTANT MESSENGERS.' Nathaniel Herbert,., 350 00 William Jackson,,.