Crustacea: Notostraca) from Serbia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Fig. Ap. 2.1. Denton Tending His Fairy Shrimp Collection
Fig. Ap. 2.1. Denton tending his fairy shrimp collection. 176 Appendix 1 Hatching and Rearing Back in the bowels of this book we noted that However, salts may leach from soils to ultimately if one takes dry soil samples from a pool basin, make the water salty, a situation which commonly preferably at its deepest point, one can then "just turns off hatching. Tap water is usually unsatis- add water and stir". In a day or two nauplii ap- factory, either because it has high TDS, or because pear if their cysts are present. O.K., so they won't it contains chlorine or chloramine, disinfectants always appear, but you get the idea. which may inhibit hatching or kill emerging If your desire is to hatch and rear fairy nauplii. shrimps the hi-tech way, you should get some As you have read time and again in Chapter 5, guidance from Brendonck et al. (1990) and temperature is an important environmental cue for Maeda-Martinez et al. (1995c). If you merely coaxing larvae from their dormant state. You can want to see what an anostracan is like, buy some guess what temperatures might need to be ap- Artemia cysts at the local aquarium shop and fol- proximated given the sample's origin. Try incu- low directions on the container. Should you wish bation at about 3-5°C if it came from the moun- to find out what's in your favorite pool, or gather tains or high desert. If from California grass- together sufficient animals for a study of behavior lands, 10° is a good level at which to start. -

Survey and Description of the Seasonal Herbaceous Wetlands (Freshwater) of the Temperate Lowland Plains in the South East of South Australia
Survey and description of the Seasonal Herbaceous Wetlands (Freshwater) of the Temperate Lowland Plains in the South East of South Australia. C.R. Dickson, L. Farrington & M. Bachmann April 2014 Report to the Department of Environment, Water and Natural Resources Page i Citation Dickson C.R., Farrington L., & Bachmann M. (2014) Survey and description of the Seasonal Herbaceous Wetlands (Freshwater) of the Temperate Lowland Plains in the South East of South Australia. Report to Department of Environment, Water and Natural Resources, Government of South Australia. Nature Glenelg Trust, Mount Gambier, South Australia. Correspondence in relation to this report contact Mr Mark Bachmann Principal Ecologist Nature Glenelg Trust (08) 8797 8181 [email protected] Cover photo: Craspedia paludicola at a Seasonal Herbaceous Wetland in Bangham Conservation Park. Disclaimer This report was commissioned by the Department of Environment, Water and Natural Resources. Although all efforts were made to ensure quality, it was based on the best information available at the time and no warranty express or implied is provided for any errors or omissions, nor in the event of its use for any other purposes or by any other parties. Page ii Acknowledgements We would like to acknowledge and thank the following people for their assistance during the project: . Private and public landholders throughout the South East of South Australia for providing access to their properties and sharing local knowledge of site history. Steve Clarke, Michael Dent, Claire Harding, and Abigail Goodman (DEWNR) and Bec Harmer (NGT) for providing field assistance on field surveys of wetlands between November 2013 and February 2014. -

2009 Annual Report Contents
EDITH COwaN UNIVERSITY School of Natural Sciences Centre for Ecosystem Management 2009 ANNUAL REPORT CONTENTS Overview 2009 3 Highlights 2009 4-7 CEM Members 8-9 Grants 10 Publications 11-13 Higher Degree Students 14 Visitors & Collaborations 15 Community Engagement Activities & Linkages 15 2 OVERVIEW 2009 The Centre for Ecosystem Management consists of 22 members from across ECU who have diverse research interests that fit into the broad themes of sustainability, biodiversity and health and ecology. In 2009 we welcomed a number of new members, mostly amongst the ranks of post doctoral and early career positions. Despite all the changes experienced with the rebirth of the CEM after the marine members split off in 2008, members had an outstanding research year. Some $0.91 million was spent on research during 2009. The magnitude of the research funding is a consequence of constructive engagement and research links between the researchers in boards or committees. The breadth and scope of the the CEM and State, National and International government contributions made by members of the CEM is remarkable organisations and research agencies. Members of the and reflects the strong research culture of the Centre and CEM also continued to produce high quality outputs in the its members. During the year 1 PhD student and 1 MSc form of refereed papers (32), reports (12) and conference student completed their studies under the supervision of proceedings (5). The breadth of research interests CEM members. in the CEM can be seen from the published outputs This report acknowledges the continued high quality and given later in this report. -

Freshwater Crustaceans As an Aboriginal Food Resource in the Northern Great Basin
UC Merced Journal of California and Great Basin Anthropology Title Freshwater Crustaceans as an Aboriginal Food Resource in the Northern Great Basin Permalink https://escholarship.org/uc/item/3w8765rq Journal Journal of California and Great Basin Anthropology, 20(1) ISSN 0191-3557 Authors Henrikson, Lael S Yohe, Robert M, II Newman, Margaret E et al. Publication Date 1998-07-01 Peer reviewed eScholarship.org Powered by the California Digital Library University of California Joumal of Califomia and Great Basin Anthropology Vol. 20, No. 1, pp. 72-87 (1998). Freshwater Crustaceans as an Aboriginal Food Resource in the Northern Great Basin LAEL SUZANN HENRIKSON, Bureau of Land Management, Shoshone District, 400 W. F Street, Shoshone, ID 83352. ROBERT M. YOHE II, Archaeological Survey of Idaho, Idaho State Historical Society, 210 Main Street, Boise, ID 83702. MARGARET E. NEWMAN, Dept. of Archaeology, University of Calgary, Alberta, Canada T2N 1N4. MARK DRUSS, Idaho Power Company, 1409 West Main Street, P.O. Box 70. Boise, ID 83707. Phyllopods of the genera Triops, Lepidums, and Branchinecta are common inhabitants of many ephemeral lakes in the American West. Tadpole shrimp (Triops spp. and Lepidums spp.) are known to have been a food source in Mexico, and fairy shrimp fBranchinecta spp.) were eaten by the aborigi nal occupants of the Great Basin. Where found, these crustaceans generally occur in numbers large enough to supply abundant calories and nutrients to humans. Several ephemeral lakes studied in the Mojave Desert arul northern Great Basin currently sustain large seasonal populations of these crusta ceans and also are surrounded by numerous small prehistoric camp sites that typically contain small artifactual assemblages consisting largely of milling implements. -

Metacommunities and Biodiversity Patterns in Mediterranean Temporary Ponds: the Role of Pond Size, Network Connectivity and Dispersal Mode
METACOMMUNITIES AND BIODIVERSITY PATTERNS IN MEDITERRANEAN TEMPORARY PONDS: THE ROLE OF POND SIZE, NETWORK CONNECTIVITY AND DISPERSAL MODE Irene Tornero Pinilla Per citar o enllaçar aquest document: Para citar o enlazar este documento: Use this url to cite or link to this publication: http://www.tdx.cat/handle/10803/670096 http://creativecommons.org/licenses/by-nc/4.0/deed.ca Aquesta obra està subjecta a una llicència Creative Commons Reconeixement- NoComercial Esta obra está bajo una licencia Creative Commons Reconocimiento-NoComercial This work is licensed under a Creative Commons Attribution-NonCommercial licence DOCTORAL THESIS Metacommunities and biodiversity patterns in Mediterranean temporary ponds: the role of pond size, network connectivity and dispersal mode Irene Tornero Pinilla 2020 DOCTORAL THESIS Metacommunities and biodiversity patterns in Mediterranean temporary ponds: the role of pond size, network connectivity and dispersal mode IRENE TORNERO PINILLA 2020 DOCTORAL PROGRAMME IN WATER SCIENCE AND TECHNOLOGY SUPERVISED BY DR DANI BOIX MASAFRET DR STÉPHANIE GASCÓN GARCIA Thesis submitted in fulfilment of the requirements to obtain the Degree of Doctor at the University of Girona Dr Dani Boix Masafret and Dr Stéphanie Gascón Garcia, from the University of Girona, DECLARE: That the thesis entitled Metacommunities and biodiversity patterns in Mediterranean temporary ponds: the role of pond size, network connectivity and dispersal mode submitted by Irene Tornero Pinilla to obtain a doctoral degree has been completed under our supervision. In witness thereof, we hereby sign this document. Dr Dani Boix Masafret Dr Stéphanie Gascón Garcia Girona, 22nd November 2019 A mi familia Caminante, son tus huellas el camino y nada más; Caminante, no hay camino, se hace camino al andar. -

VERNAL POOL TADPOLE SHRIMP Lepidurus Packardi
U.S. Fish & Wildlife Service Sacramento Fish & Wildlife Office Species Account VERNAL POOL TADPOLE SHRIMP Lepidurus packardi CLASSIFICATION: Endangered Federal Register 59-48136; September 19, 1994 http://ecos.fws.gov/docs/federal_register/fr2692.pdf On October 9, 2007, we published a 5-year review recommending that the species remain listed as endangered. CRITICAL HABITAT: Designated Originally designated in Federal Register 68:46683; August 6, 2003. The designation was revised in FR 70:46923; August 11, 2005. Species by unit designations were published in FR 71:7117 (PDF), February 10, 2006. RECOVERY PLAN: Final Recovery Plan for Vernal Pool Ecosystems of California and Southern Oregon, December 15, 2005. http://ecos.fws.gov/docs/recovery_plan/060614.pdf DESCRIPTION The vernal pool tadpole shrimp (Lepidurus packardi) is a small crustacean in the Triopsidae family. It has compound eyes, a large shield-like carapace (shell) that covers most of the body, and a pair of long cercopods (appendages) at the end of the last abdominal segment. Vernal pool tadpole shrimp adults reach a length of 2 inches in length. They have about 35 pairs of legs and two long cercopods. This species superficially resembles the rice field tadpole shrimp (Triops longicaudatus). Tadpole shrimp climb or scramble over objects, as well as plowing along or within bottom sediments. Their diet consists of organic debris and living organisms, such as fairy shrimp and other invertebrates. This animal inhabits vernal pools containing clear to highly turbid water, ranging in size from 54 square feet in the former Mather Air Force Base area of Sacramento County, to the 89-acre Olcott Lake at Jepson Prairie. -

Living Fossils
Living Fossils 1. Diplopanax - Creation Ex Nihilo 12(4): 6,7 (Sept - Nov 1990) 2. Tuatara - Biblical Basis for Modern Science by Henry Morris (Baker Book House, Grand Rapids, Michigan, 1984) pg.355; Scientific Creationism Edited by Henry Morris (Master Books: El Cajon, CA, 1974) pg.89; The Genesis Flood by John Whitcomb & Henry Morris (Presbyterian & Reformed Publishing: Philipsburg, NJ, 1961) pgs.176, 177; The Creation-Evolution Controversy by R.L. Wysong (Inquiry Press: Midland, Michigan, 1976) pg.287, 289; "The Tuatara: Why is it a lone survivor?" by C. M. Bogert, Scientific Monthly, 76 (1953): 165; Sphenodon - Gliedman "Miracle Mutations", Science Digest (Feb, 1982) pgs.90, 92; A Case for Creation by Wayne Frair & Percival Davis (Moody Press, 1967) pg.65 3. Latimeria chalumnae (Coelacanth) - Also mentioned in Creation Ex Nihilo 15(4): 45 (Sept - Nov, 1993); Creation 23(2): 5 (March - May, 2001); Forey, "The Coelacanth as a Living Fossil" in Living Fossils, N. Eldredge & S. Stanley, eds, 1984) pg.166; A Case for Creation by Wayne Frair & Percival Davis (Moody Press, 1967) pg.65; Genes, Genesis & Evolution by John W. Klotz (Concordia Publishing House: St. Louis, Missouri, 1955) pgs.200-202; Darwin Retried: an appeal to reason by Norman Macbeth (Harvard Common Press: Boston, Massachusetts, 1971) pg.121; Biology: A Search for Order in Complexity, Edited by John N. Moore & Harold Slusher (Zondervan Publishing House: Grand Rapids, Michigan, 1970) pg.264; The Biblical Basis for Modern Science by Henry M. Morris (Baker Book House, Grand Rapids, Michigan, 1984) pg.355; Scientific Creationism Edited by Henry Morris (Master Books: El Cajon, CA, 1974) pg.89; The Genesis Flood by John Whitcomb & Henry Morris (Presbyterian & Reformed Publishing: Philipsburg, NJ, 1961) pgs.177,178; After Its Kind by Byron C. -

Crustacea, Notostraca) from Iran
Journal of Biological Research -Thessaloniki 19 : 75 – 79 , 20 13 J. Biol. Res. -Thessalon. is available online at http://www.jbr.gr Indexed in: Wos (Web of science, IsI thomson), sCOPUs, CAs (Chemical Abstracts service) and dOAj (directory of Open Access journals) — Short CommuniCation — on the occurrence of Lepidurus apus (Linnaeus, 1758) (Crustacea, notostraca) from iran Behroz AtAshBAr 1,2 *, N aser Agh 2, L ynda BeLAdjAL 1 and johan MerteNs 1 1 Department of Biology , Faculty of Science , Ghent University , K.L. Ledeganckstraat 35 , 9000 Gent , Belgium 2 Department of Biology and Aquaculture , Artemia and Aquatic Animals Research Institute , Urmia University , Urmia-57153 , Iran received: 16 November 2011 Accepted after revision: 18 july 2012 the occurrence of Lepidurus apus in Iran is reported for the first time. this species was found in Aigher goli located in the mountains area, east Azerbaijan province (North east, Iran). details on biogeography, ecology and morphology of this species are provided. Key words: Lepidurus apus , Aigher goli, east Azerbaijan, Iran. INtrOdUCtION their world-wide distribution is due to their anti qu - ity, but possibly also to their passive transport: geo - Notostracan records date back to the Carboniferous graphical barriers are more effective for non-passive - and possibly up to the devonian period (Wallossek, ly distributed animals. From an ecological point of 1993, 1995; Kelber, 1998). In fact, there are Upper view, notostracans, like most branchiopods, are re - triassic Triops fossils from germany -

Lepidurus Arcticus (Crustacea : Notostraca); an Unexpected Prey of Arctic Charr (Salvelinus Alpinus) in a High Arctic River
BOREAL ENVIRONMENT RESEARCH 23: 149–157 © 2018 ISSN 1797-2469 (online) Helsinki 20 April 2018 Lepidurus arcticus (Crustacea : Notostraca); an unexpected prey of Arctic charr (Salvelinus alpinus) in a High Arctic river Reidar Borgstrøm1)*, Morten Aas1)2), Hanne Hegseth1)2), J. Brian Dempson3) and Martin-Arne Svenning2) 1) Faculty of Environmental Sciences and Natural Resource Management, Norwegian University of Life Sciences, P.O. Box 5003, NO-1432 Ås, Norway (*corresponding author’s e-mail: reidar.borgstrom@ nmbu.no) 2) Norwegian Institute of Nature Research, Arctic Ecology Department, Fram Center, P.O. Box 6606 Langnes, NO-9296 Tromsø, Norway 3) Fisheries & Oceans Canada, Science Branch, 80 East White Hills Road, P.O. Box 5667, St. John’s, NL A1C 5X1, Canada Received 20 Dec. 2017, final version received 16 Mar. 2018, accepted 12 Apr. 2018 Borgstrøm R., Aas M., Hegseth H., Dempson J.B. & Svenning M.-A. 2018: Lepidurus arcticus (Crusta- cea : Notostraca); an unexpected prey of Arctic charr (Salvelinus alpinus) in a High Arctic river. Boreal Env. Res. 23: 149–157. The phyllopod Lepidurus arcticus, commonly called the arctic tadpole shrimp, is an important food item of both Arctic charr Salvelinus alpinus and brown trout Salmo trutta in lakes in Iceland and Scandinavia, especially at low fish densities. In the High Arctic, the tadpole shrimp is abundant in fishless localities, but absent or rare in most lakes where fish are present. We studied the diet of Arctic charr in the Straumsjøen watercourse on Spitsbergen, the main island of Svalbard. The outlet river is dry nine months of the year, and all Arctic charr present during summer have descended from the lake. -

Crustacean Phylogeny…? Nauplius • First Larva Stage of Most “It Can Be Concluded That Crustacean Crustaceans
Bio 370 Crustacea Main arthropod clades (Regier et al 2010) Phylum Arthropoda http://blogs.discoverm • Trilobita agazine.com/loom/201 0/02/10/blind-cousins- Subphylum (or Class) Crustacea to-the-arthropod- • Chelicerata superstars/ Mostly aquatic, with calcified exoskeleton. • Mandibulata – Myriapoda (Chilopoda, Diplopoda) Head derived from acron plus next five segments- so primitively has 5 pairs of appendages: – Pancrustacea • Oligostraca (Ostracoda, Branchiura) -2 pair antennae • Altocrustacea - 1 pair of jaws – Vericrustacea - 2 pair of maxillae » (Branchiopoda, Decapoda) - usually a median (cyclopean) eye and – Miracrustacea one pair of compound eyes » Xenocarida (Remipedia, Cephalocarida) » Hexapoda Tagmosis of trunk varies in different taxa Crustacean phylogeny…? Nauplius • first larva stage of most “It can be concluded that crustacean crustaceans. phylogeny remains essentially unresolved. • three pairs of appendages • single median (naupliar) eye Conflict is rife, irrespective of whether one compares different morphological studies, molecular studies, or both.” Appendages: Jenner, 2010: Arthropod Structure & Development 39:143– -1st antennae 153 -2nd antennae - mandibles 1 Bio 370 Crustacea Crustacean taxa you should know Remipede habitat: a sea cave “blue hole” on Andros Island. Seven species are found in the Bahamas. Class Remipedia Class Malacostraca Class Branchiopoda “Peracarida”-marsupial crustacea Notostraca –tadpole shrimp Isopoda- isopods Anostraca-fairy shrimp Amphipoda- amphipods Cladocera- water fleas Mysidacea- mysids Conchostraca- clam shrimp “Eucarida” Class Maxillopoda Euphausiacea- krill Ostracoda- ostracods Decapoda- decapods- ten leggers Copepoda- copepods Branchiura- fish lice Penaeoidea- penaeid shrimp Cirripedia- barnacles Caridea- carid shrimp Astacidea- crayfish & lobsters Brachyura- true crabs Anomura- false crabs “Stomatopoda”– mantis shrimps Class Remipedia Remipides found only in sea caves in the Caribbean, the Canary Islands, and Western Australia (see pink below). -
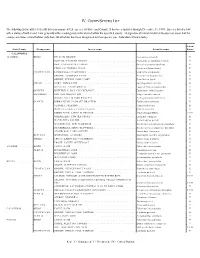
Iv. County/Species List
IV. COUNTY/SPECIES LIST The following list identifies federally listed or proposed U.S. species by State and County. It has been updated through December 31, 1999. Species listed below with a status of both E and T are generally either endangered or threatened within the specified county. Designation of critical habitat (CH) does not mean that the county constitutes critical habitat, only that critical habitat has been designated for that species (see Addendum A Instructions). Action/ State/County Group name Inverse name Scientific name Status CALIFORNIA ALAMEDA ......... BIRDS ....... PELICAN, BROWN ........................................ Pelicanus occidentalis E PLOVER, WESTERN SNOWY ................................ Charadrius alexandrinus nivosus T RAIL, CALIFORNIA CLAPPER ............................... Rallus longirostris obsoletus E TERN, CALIFORNIA LEAST ................................. Sterna antillarum browni E CRUSTACEAN . LINDERIELLA, CALIFORNIA ................................ Linderiella occidentalis E SHRIMP, LONGHORN FAIRY ................................ Branchinecta longiantenna E SHRIMP, VERNAL POOL FAIRY .............................. Branchinecta lynchi T FISHES ...... GOBY, TIDEWATER ...................................... Eucyclogobius newberryi E SPLITTAIL, SACRAMENTO ................................. Pogonichthys macrolepidotus T INSECTS ..... BUTTERFLY, BAY CHECKERSPOT ........................... Euphydryas editha bayensis T MAMMALS ... FOX, SAN JOAQUIN KIT .................................... Vulpes macrotis -

On Homology of Arthropod Compound Eyes' "The Eye" Has Long Served As
INTEGR. CotoP. BIOL., 43:522-530 (2003) On Homology of Arthropod Compound Eyes' TODD H. OAKLEY 2 Ecology Evolutioni and Marine Biology, University of California-SantaBarbara, S'anzta Barbara, Califonzia 93106 SyNopsis. Eyes serve as models to understand the evolution of complex traits, with broad implications for the origins of evolutionary novelty. Discussions of eye evolution are relevant at miany taxonomic levels, especially within arthropods where compound eye distribution is perplexing. Either compound eyes were lost numerous times or very similar eyes evolved separately in multiple lineages. Arthropod compound eye homology is possible, especially; between crustaceans and hexapods, which have very similar eye facets and may be sister taxa. However, judging homology only on similarity requires subjective decisions. Regardless of whether compound eyes were present in a common ancestor of arthropods or crustaceans + hexapods, recent phylogenetic evidence suggests that the compound eyes, today present in myodocopid ostracods (Crus- tacea), may have been absent in ostracod ancestors. This pattern is inconsistent with phylogenetic homology. Multiple losses of ostracod eyes are an alternative hypothesis that is statistically improbable and without clear cause. One possible evolutionary process to explain the lack of phylogenetic'homology of ostracod compound eyes is that eyes may evolve by switchback evolution, where genes for lost structures remain dormant and are re-expressed much later in evolution. INTRODUCTION 'the recent evidence for and implications of a poten- "The eye" has long served as a canonical example tially non-homologous arthropod compound eye, un- of a complex trait. In an early design-based argument derstanding the case for compound eye homology is for the existence of God, Paley (1846) used the eye as important.