Relating the 4-Year Lemming Population Cycle to a 3.8-Year Lunar Cycle and ENSO
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Long-Term Monitoring Reveals Topographical Features and Vegetation Explain Winter Habitat Use of an Arctic Rodent
bioRxiv preprint doi: https://doi.org/10.1101/2021.01.24.427984; this version posted January 26, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 1 Title 2 Long-term monitoring reveals topographical features and vegetation explain winter habitat 3 use of an Arctic rodent 4 Abstract 5 Collapsing lemming cycles have been observed across the Arctic, presumably due to global 6 warming creating less favorable winter conditions. The quality of wintering habitats, such as 7 depth of snow cover, plays a key role in sustaining population dynamics of arctic lemmings. 8 However, few studies so far investigated habitat use during the arctic winter. Here, we used a 9 unique long-term time series to test whether lemmings are associated with topographical and 10 vegetational habitat features for their winter refugi. We examined yearly numbers and 11 distribution of 22,769 winter nests of the collared lemming Dicrostonyx groenlandicus from 12 an ongoing long-term research on Traill Island, Northeast Greenland, collected between 1989 13 and 2019, and correlated this information with data on dominant vegetation types, elevation 14 and slope. We specifically asked if lemming nests were more frequent at sites with preferred 15 food plants such as Dryas octopetala x integrifolia and at sites with increased snow cover. We 16 found that the number of lemming nests was highest in areas with a high proportion of Dryas 17 heath, but also correlated with other vegetation types which suggest some flexibility in 18 resource use of wintering lemmings. -

Mammal Extinction Facilitated Biome Shift and Human Population Change During the Last Glacial Termination in East-Central Europeenikő
Mammal Extinction Facilitated Biome Shift and Human Population Change During the Last Glacial Termination in East-Central EuropeEnikő Enikő Magyari ( [email protected] ) Eötvös Loránd University Mihály Gasparik Hungarian Natural History Museum István Major Hungarian Academy of Science György Lengyel University of Miskolc Ilona Pál Hungarian Academy of Science Attila Virág MTA-MTM-ELTE Research Group for Palaeontology János Korponai University of Public Service Zoltán Szabó Eötvös Loránd University Piroska Pazonyi MTA-MTM-ELTE Research Group for Palaeontology Research Article Keywords: megafauna, extinction, vegetation dynamics, biome, climate change, biodiversity change, Epigravettian, late glacial Posted Date: August 11th, 2021 DOI: https://doi.org/10.21203/rs.3.rs-778658/v1 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 1/27 Abstract Studying local extinction times, associated environmental and human population changes during the last glacial termination provides insights into the causes of mega- and microfauna extinctions. In East-Central (EC) Europe, Palaeolithic human groups were present throughout the last glacial maximum (LGM), but disappeared suddenly around 15 200 cal yr BP. In this study we use radiocarbon dated cave sediment proles and a large set of direct AMS 14C dates on mammal bones to determine local extinction times that are compared with the Epigravettian population decline, quantitative climate models, pollen and plant macrofossil inferred climate and biome reconstructions and coprophilous fungi derived total megafauna change for EC Europe. Our results suggest that the population size of large herbivores decreased in the area after 17 700 cal yr BP, when temperate tree abundance and warm continental steppe cover both increased in the lowlands Boreal forest expansion took place around 16 200 cal yr BP. -

The Role of Diseases in Mass Mortality of Wood Lemmings (Myopus Schisticolor)
The role of diseases in mass mortality of wood lemmings (Myopus schisticolor) Sjukdomars roll i massutdöende av skogslämmel (Myopus schisticolor) Henrik Johansen Master’s thesis • 30 credits Swedish University of Agricultural Sciences, SLU Department of Wildlife, Fish, and Environmental Studies Forest Science programme Examensarbete/Master’s thesis, 2021:7 Umeå, Sweden 2021 The role of disease in mass mortality of wood lemming (Myopus schisticolor) Sjukdomars roll I massutdöende av skogslämmel (Myopus schisticolor) Henrik Johansen Supervisor: Frauke Ecke, Swedish University of Agricultural Science, Department of wildlife, Fish, and Environmental Studies Assistant supervisor: Magnus Magnusson, Swedish University of Agricultural Science, Department of wildlife, Fish, and Environmental Studies Examiner: Joris Cromsigt, Swedish University of Agricultural Science, Department of wildlife, Fish, and Environmental Studies Credits: 30 credits Level: Second cycle, A2E Course title: Master’s thesis in Forest Science, A2E - Wildlife, Fish, and Environmental Studies Course code: EX0840 Programme/education: Forest Science programme Course coordinating dept: Department of Wildlife, Fish, and Environmental Studies Place of publication: Umeå, Sweden Year of publication: 2021 Cover picture: Thomas Secher Jensen Title of series: Examensarbete/Master’s thesis Part number: 2021:7 Keywords: Wood lemming, Myopus schisticolor, Disease, Virus, Pathogens, Mass mortality, Orthohantavirus, Pan-orthohantavirus, Somatic index, Spleen index Swedish University of Agricultural Sciences Faculty of Forest Science Department of Wildlife, Fish, and Environmental Studies Publishing and archiving Approved students’ theses at SLU are published electronically. As a student, you have the copyright to your own work and need to approve the electronic publishing. If you check the box for YES, the full text (pdf file) and metadata will be visible and searchable online. -

DNA Metabarcoding of Stomach Contents
1 Shedding new light on the diet of Norwegian lemmings: DNA 2 metabarcoding of stomach content 3 4 Eeva M. Soininen 1*, Lucie Zinger 2,7 , Ludovic Gielly 2, Eva Bellemain 3,6 , Kari Anne Bråthen 1, 5 Christian Brochmann 3, Laura S. Epp 3, Galina Gussarova 3, Kristian Hassel 4, John-André 6 Henden 1, Siw T. Killengreen 1, Teppo Rämä 5, Hans K. Stenøien 4, Nigel G. Yoccoz 1, Rolf A. Ims 1 7 1 Department of Arctic and Marine Biology, University of Tromsø, NO-9037 Tromsø, Norway 8 2Laboratoire d’ECologie Alpine, UMR UJF-UdS-CNRS 5553, Université Joseph Fourier, 2233 9 rue de la Piscine, BP 53, 38041 Grenoble Cedex 9, France 10 3 National Centre for Biosystematics, Natural History Museum, University of Oslo, PO 11 Box 1172 Blindern, NO-0318 Oslo, Norway 12 4 Museum of Natural History, Norwegian University of Science and Technology, NO-7491 13 Trondheim, Norway 14 5 Tromsø University Museum, University of Tromsø, NO-9037 Tromsø, Norway 15 6Current address: Savoie Technolac, Batiment House Boat n7 12, allee du lac de Garde, BP 16 274, 73375 Le Bourget-du-Lac Cedex, France 17 7Current address: Laboratoire Evolution et Biodiversité Biologique, UMR 5174, Université 18 Paul Sabatier, 118 route de Narbonne, 31062 Toulouse Cedex 9, France 19 *Corresponding author e-mail: [email protected] , tel: +47 77620932, 20 fax: +47 77 64 63 33 1 21 Abstract 22 Lemmings are key herbivores in many arctic food webs and their population dynamics have 23 major impacts on the functioning of tundra systems. -

Prey Selection and Prey Handling in Two Raptors During the Breeding Season As Revealed by the Use of Video Monitoring
Norwegian University of Life Sciences Department of Ecology and Natural Resource Management (INA) Master Thesis 2015 30 credits Prey selection and prey handling in two raptors during the breeding season as revealed by the use of video monitoring Byttedyrvalg og byttedyrhåndtering hos to rovfugler i hekkeperioden, avslørt ved bruk av videoovervåking Ida Dihle A C K N O W L E D G E M E N T S First of all, I would like to thank Fylkesmannen i Oppland for the financial support. I would like to give a huge thanks to my brilliant supervisor Geir A. Sonerud who has spend hours identifying prey, helping with the statistical analyses and thoroughly reviewing my drafts. I am also very grateful to supervisor Ronny Steen, for all his effort regarding all stages of the fieldwork, his technical support throughout the process and providing helpful comments on my drafts. I would like to direct a big thanks to Vidar Selås, who contributed with his expertise when identifying prey. I am very grateful for the effort, inspiration and company during fieldwork given by Helge Grønnlien. The last weeks of recordings was made possible because of him. Dag S. Rusten also deserves a thanks, who along with Ronny Steen and Helge Grønnlien installed the cameras at the nests. My friends, Lene M. Rabben in particular, and family has been a big help throughout the process. I would like to thank my parents for providing equipment, and joining me in field, and Thea Dihle for punching numbers in the less inspiring periods of the writing process. -

Sex Chromosome Translocations
RESEARCH ARTICLE Rapid Karyotype Evolution in Lasiopodomys Involved at Least Two Autosome ± Sex Chromosome Translocations Olga L. Gladkikh1☯, Svetlana A. Romanenko1,2☯*, Natalya A. Lemskaya1, Natalya A. Serdyukova1, Patricia C. M. O'Brien3, Julia M. Kovalskaya4, Antonina V. Smorkatcheva5, Feodor N. Golenishchev6, Polina L. Perelman1,2, Vladimir A. Trifonov1,2, Malcolm A. Ferguson-Smith3, Fengtang Yang7, Alexander S. Graphodatsky1,2 a11111 1 Institute of Molecular and Cellular Biology, Siberian Branch of the Russian Academy of Sciences, Novosibirsk, Russia, 2 Novosibirsk State University, Novosibirsk, Russia, 3 Cambridge Resource Centre for Comparative Genomics, Department of Veterinary Medicine, University of Cambridge, Cambridge, United Kingdom, 4 Severtzov Institute of Ecology and Evolution, Russian Academy of Sciences, Moscow, Russia, 5 Department of Vertebrate Zoology, Saint Petersburg State University, Saint Petersburg, Russia, 6 Zoological Institute, Russian Academy of Sciences, Saint Petersburg, Russia, 7 Wellcome Trust Sanger Institute, Wellcome Genome Campus, Hinxton, Cambridge, United Kingdom ☯ These authors contributed equally to this work. OPEN ACCESS * [email protected] Citation: Gladkikh OL, Romanenko SA, Lemskaya NA, Serdyukova NA, O'Brien PCM, Kovalskaya JM, et al. (2016) Rapid Karyotype Evolution in Abstract Lasiopodomys Involved at Least Two Autosome ± Sex Chromosome Translocations. PLoS ONE 11 The generic status of Lasiopodomys and its division into subgenera Lasiopodomys (L. man- (12): e0167653. doi:10.1371/journal. pone.0167653 darinus, L. brandtii) and Stenocranius (L. gregalis, L. raddei) are not generally accepted because of contradictions between the morphological and molecular data. To obtain cyto- Editor: Igor V. Sharakhov, Virginia Tech, UNITED STATES genetic evidence for the Lasiopodomys genus and its subgenera and to test the autosome to sex chromosome translocation hypothesis of sex chromosome complex origin in L. -

1 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 This Paper Explores
Small mammals and paleovironmental context of the terminal pleistocene and early holocene human occupation of central Alaska Item Type Article Authors Lanoë, François B.; Reuther, Joshua D.; Holmes, Charles E.; Potter, Ben A. Citation Lanoë, FB, Reuther, JD, Holmes, CE, Potter, BA. Small mammals and paleovironmental context of the terminal pleistocene and early holocene human occupation of central Alaska. Geoarchaeology. 2019; 1– 13. https://doi.org/10.1002/gea.21768 DOI 10.1002/gea.21768 Publisher WILEY Journal GEOARCHAEOLOGY-AN INTERNATIONAL JOURNAL Rights © 2019 Wiley Periodicals, Inc. Download date 07/10/2021 13:55:14 Item License http://rightsstatements.org/vocab/InC/1.0/ Version Final accepted manuscript Link to Item http://hdl.handle.net/10150/634952 Page 2 of 42 1 2 3 1 SMALL MAMMALS AND PALEOVIRONMENTAL CONTEXT OF THE TERMINAL 4 5 2 PLEISTOCENE AND EARLY HOLOCENE HUMAN OCCUPATION OF CENTRAL 6 3 ALASKA 7 8 4 François B. Lanoëab, Joshua D. Reutherbc, Charles E. Holmesc, and Ben A. Potterc 9 10 5 11 a 12 6 Bureau of Applied Research in Anthropology, University of Arizona, 1009 E S Campus Dr, 13 7 Tucson, AZ 85721 14 bArchaeology Department, University of Alaska Museum of the North, 1962 Yukon Dr, 15 8 16 9 Fairbanks, AK 99775 17 10 cDepartment of Anthropology, University of Alaska, 303 Tanana Loop, Fairbanks, AK 99775 18 19 11 20 21 12 Corresponding author: François Lanoë, [email protected] 22 23 13 24 25 14 Abstract 26 27 15 This paper explores paleoenvironmental and paleoecological information that may be obtained 28 16 from small-mammal assemblages recovered at central Alaska archaeological sites dated to the 29 30 17 Terminal Pleistocene and Early Holocene (14,500-8000 cal B.P.). -

A-Nadachowski.Vp:Corelventura
Acta zoologica cracoviensia, 58(2): 181-194, Kraków, 31 December, 2015 Ó Institute of Systematics and Evolution of Animals, Pol. Acad. Sci., Kraków doi:10.3409/azc.58_2.181 NewLatePleistocenefaunalassemblagesfromPodhaleBasin, WesternCarpathians,Poland:preliminaryresults AdamNADACHOWSKI and Pawe³ VALDE-NOWAK Received: 05 October 2015. Accepted: 21 December 2015. Available online: 30 December 2015. NADACHOWSKI A., VALDE-NOWAK P. 2015. New Late Pleistocene faunal assem- blages from Podhale Basin, Western Carpathians, Poland: preliminary results. Acta zool. cracov., 58(2): 181-194. Abstract. Interdisciplinary studies in Ob³azowa Cave undertaken near its western entrance and in a new archaeological site in Cisowa Rock yielded finely layered sedimentary se- quences with an abundant fossil fauna of mainly small vertebrates and scarce archaeologi- cal finds. In this paper we present a preliminary palaeoenvironmental and palaeoclimatic reconstruction of the sequences dated around MIS 2/MIS 1 boundary on the basis of small mammal assemblages. The faunal changes appear to be gradual in spite of abrupt climatic changes expected to occur at that time. The environmental and faunal results are discussed with other sequences of similar age in Poland. Key words: fossil fauna, habitat and climate change, steppe-tundra, radiocarbon dating, Ob³azowa Cave. * Adam NADACHOWSKI, Institute of Systematics and Evolution of Animals Polish Acad- emy of Sciences, S³awkowska Str. 17, 31-016 Kraków, Poland. E-mail: [email protected] Pawe³ VALDE-NOWAK, Institute of Archaeology, Jagiellonian University, Go³êbia Str. 11, 31-007 Kraków, Poland. E-mail: [email protected] I.INTRODUCTION The Podhale Basin, also known as Orawa-Nowy Targ Basin, is located in the Western Carpathians, south Poland. -

Roosevelt Wild Life Bulletins the Roosevelt Wild Life Station
SUNY College of Environmental Science and Forestry Digital Commons @ ESF Roosevelt Wild Life Bulletins The Roosevelt Wild Life Station 1930 Roosevelt Wild Life Bulletin Charles E. Johnson SUNY College of Environmental Science and Forestry Follow this and additional works at: https://digitalcommons.esf.edu/rwlsbulletin Part of the Animal Sciences Commons, Biodiversity Commons, Ecology and Evolutionary Biology Commons, and the Natural Resources and Conservation Commons Recommended Citation Johnson, Charles E., "Roosevelt Wild Life Bulletin" (1930). Roosevelt Wild Life Bulletins. 16. https://digitalcommons.esf.edu/rwlsbulletin/16 This Book is brought to you for free and open access by the The Roosevelt Wild Life Station at Digital Commons @ ESF. It has been accepted for inclusion in Roosevelt Wild Life Bulletins by an authorized administrator of Digital Commons @ ESF. For more information, please contact [email protected], [email protected]. VOL. Ill AUGUST. 1930 No. 2c BULLETIN OF The New York State College of Forestry AT Syracuse University HUGH P. BAKER, Dean Roosevelt Wild Life Bulletin VOLUME 5 NUMBER 4 OF THE Roosevelt Wild Life Forest Experiment Station '-THE BIOLOGY OF THE VOLES OF NEW YORK / - ' jHE RELATION OF MAMMALS TO THE HARVARD FOREST CONTENTS OF RECENT ROOSEVELT WILD LIFE BULLETINS (To obtain these publications see announcement on back of title page.) Roosevelt Wild Life Bulletin, Vol. i, No. 3. March, 1923. 1. The Summer Birds of the Allegany State Park. .Aretas A. Saunders. 2. The Ruffed Grouse, with Special Reference to its Drumming Edmund J. Sawyer. 3. Current Station Notes The Director and Editor. Roosevelt Wild Life Bulletin, Vol. i. No. -

Norway's Environmental Targets
Norway’s Environmental Targets Action for the environment 3 ————————— Priority areas and tools Useful for achieving our goals 4 websites: Priority areas www.miljo.no 01 Living seas and coastal environments 6 www.environment.no 02 Healthy lakes and rivers 10 www.klif.no 03 Rich and varied wetlands 14 www.dirnat.no 04 Forest diversity 18 www.npolar.no 05 Spectacular mountain landscapes 22 www.ra.no 06 A valuable cultural heritage 26 www.statkart.no 07 A good urban environment 30 08 The great outdoors 34 ————————— 09 An unpolluted environment 38 10 Clean air 42 11 A stable climate 46 Instruments Knowledge and geographical information 50 Legislation and planning procedures 52 International cooperation 54 Norway’s Environmental Targets was published in Norwegian in 2011. This translated version contains some updates and corrections, but has not been completely revised. 7U\NN$QGYRUG*UDILVN$6 Action for the environment Thirty years ago, acid rain was killing fish People have always left evidence of their in Norwegian rivers. Now, both salmon and activities behind them, altering the environ- trout are back again in many rivers thanks to ment for better or worse. Our cultural herit- concerted national and international efforts age does more than tell our history – it is also to cut emissions. part of a dynamic contemporary environ- ment. But it, too, is vulnerable: once build- Norway’s nature reaches from the deepest ings and cultural landscapes are destroyed seabed to the topmost mountain peaks, or removed, they are lost for ever. and from the thickest forest to the highest waterfall. -
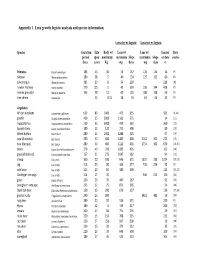
Appendix 1. Lens Growth Logistic Analysis and Species Information
Appendix 1. Lens growth logistic analysis and species information. Lens dry wt logistic Lens wet wt logistic Species Gestation Life Body wt Lens wt Lens wt Lenses Data period span maximum maximum Slope maximum Slope or data source days years Kg mg days mg days n Primates Papio hamadryas 183 45 30 55 167 178 124 16 PS baboon Macaca fascicularis 180 38 5 40 130 125 162 60 PS cynomolgus Alouatta caraya 185 20 8 34 220 228 [8] howler monkey Homo sapiens 270 115 5 45 359 198 244 608 PS human prenatal Macaca mulatta 165 38 12 60 125 188 108 50 PS tree shrew Tupaia glis 12 0.15 28 76 63 63 32 PS Ungulates African elephant Loxodonta a africana 659 80 5000 475 935 561 [9,10] giraffe Giraffa camelopardalis 460 35 2000 1163 571 34 [11] hipppotamus Hippopotamus amphibius 250 55 3000 410 362 360 [12] Spanish Ibex Capra pyrenaica Schinz 180 12 120 711 508 80 [13] Wood buffalo Bison bison 280 25 1000 1288 525 97 [14] cow (Australia) Bos taurus 280 30 600 1483 400 3162 302 231 [15] cow (Europe) Bos taurus 280 30 400 1122 401 2754 302 630 [16,17] zebra Equus burchelli antiquorum 370 40 350 1085 436 102 [18] gnu (wildebeest) Connochaetes taurinus 257 21 275 1047 362 94 [19] sheep Ovis aries 160 20 100 646 311 1622 239 1224 [20,21] pig Sus scrofa 115 25 80 364 277 736 254 92 PS wild boar Sus scrofa 115 20 84 389 309 113 [22,23] Gottingen minipigs Sus scrofa 114 17 35 646 211 100 [24] goat Capra hircus L 150 20 70 447 267 91 [25] pronghorn antelope Antilocapra americana 235 12 75 871 365 24 [26] black tail deer Odocoileus hemionus columbianus 150 15 200 -

Ice Age Animal Factsheets
Ice Age Animal Factsheets Scientists have found frozen woolly mammoths in Height: 3m (shoulder) cold places, like Russia. Weight: 6000 kg Some of these mammoths Diet: Herbivore were so complete that you Range: Eurasia & can see their hair and N. America muscles! WOOLLY MAMMOTH HERBIVORES: MEGAFAUNA The word “megafauna” means “big animal”. Many animals in the Ice Age were very large compared with creatures living today. WOOLLY RHINOCEROS Woolly rhinos were plant- eaters. They used their Height: 2m (shoulder) massive horns to scrape Weight: 2700 kg away ice and snow from Diet: Herbivore plants so they could feed Range: Eurasia (like a giant snow plough)! In Spain, there is a cave called Altamira which has Height: 2m (shoulder) beautiful Ice Age paintings Weight: 900 kg of bison. We also have Diet: Herbivore an Ice Age carv ing of a Range: Eurasia bison at Creswell Crags! STEPPE BISON HERBIVORES: COW-LIKE ANIMALS Aurochs and bison were large animals which looked a lot like bulls. Humans during the Ice Age would have hunted them for their meat and skins. Bison were one of the most popular The farm cows we have choicesAUROCHS of subject in Stone Age art. today are related to the Height: 1.8m (shoulder) aurochs. Scientists think Weight: 700 kg humans began to tame Diet: Herbivore aurochs around ten thousand Range: Eurasia & years ago. Africa The saiga antelope has a very unusual nose which Height: 0.7m (shoulder) helps them live in cold places. Breathing Weight: 40 kg through their nose lets Diet: Herbivore them warm up cold air! Range: Eurasia SAIGA ANTELOPE HERBIVORES: GRAZERS Grazers are animals which eat food on the ground, usually grasses or herby plants.