Exosomal Long Non-Coding RNA LINC00662 Promotes Non-Small Cell Lung Cancer Progression by Mir-320D/E2F1 Axis
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Annual Report Annual Report 2020
2020 Annual Report Annual Report 2020 For further details about information disclosure, please visit the website of Yanzhou Coal Mining Company Limited at Important Notice The Board, Supervisory Committee and the Directors, Supervisors and senior management of the Company warrant the authenticity, accuracy and completeness of the information contained in the annual report and there are no misrepresentations, misleading statements contained in or material omissions from the annual report for which they shall assume joint and several responsibilities. The 2020 Annual Report of Yanzhou Coal Mining Company Limited has been approved by the eleventh meeting of the eighth session of the Board. All ten Directors of quorum attended the meeting. SHINEWING (HK) CPA Limited issued the standard independent auditor report with clean opinion for the Company. Mr. Li Xiyong, Chairman of the Board, Mr. Zhao Qingchun, Chief Financial Officer, and Mr. Xu Jian, head of Finance Management Department, hereby warrant the authenticity, accuracy and completeness of the financial statements contained in this annual report. The Board of the Company proposed to distribute a cash dividend of RMB10.00 per ten shares (tax inclusive) for the year of 2020 based on the number of shares on the record date of the dividend and equity distribution. The forward-looking statements contained in this annual report regarding the Company’s future plans do not constitute any substantive commitment to investors and investors are reminded of the investment risks. There was no appropriation of funds of the Company by the Controlling Shareholder or its related parties for non-operational activities. There were no guarantees granted to external parties by the Company without complying with the prescribed decision-making procedures. -

Reliability Optimization of a Railway Network
sustainability Article Reliability Optimization of a Railway Network Xuelei Meng 1,2,*, Yahui Wang 3, Limin Jia 2 and Lei Li 4 1 School of Traffic and Transportation, Lanzhou Jiaotong University, Lanzhou 730070, Gansu, China 2 State Key Laboratory of Rail Traffic Control and Safety, Beijing Jiaotong University, Beijing 100044, China; [email protected] 3 School of Foreign Languages, Lanzhou Jiaotong University, Lanzhou 730070, Gansu, China; [email protected] 4 Key Laboratory of Urban Rail Transit Intelligent Operation and Maintenance Technology & Equipment of Zhejiang Province, Zhejiang Normal University, Jinhua 321004, Zhejiang, China; [email protected] * Correspondence: [email protected] Received: 7 October 2020; Accepted: 14 November 2020; Published: 24 November 2020 Abstract: With the increase of the railway operating mileage, the railway network is becoming more and more complicated. We expect to build more railway lines to offer the possibility to offer more high quality service for the passengers, while the investment is often limited. Therefore, it is very important to decide the pairs of cities to add new railway lines under the condition of limited construction investment in order to optimize the railway line network to maximize the reliability of the railway network to deal with the railway passenger transport task under emergency conditions. In this paper, we firstly define the reliability of the railway networks based on probability theory by analyzing three minor cases. Then we construct a reliability optimization model for the railway network to solve the problem, expecting to enhance the railway network with the limited investment. The goal is to make an optimal decision when choosing where to add new railway lines to maximize the reliability of the whole railway network, taking the construction investment as the main constraint, which is turned to the building mileage limit. -

Supply Chain Strategic Alliances Partner Selection for Rizhao Port
International Journal of Scientific and Research Publications, Volume 4, Issue 11, November 2014 1 ISSN 2250-3153 Supply Chain Strategic Alliances Partner Selection for Rizhao Port Libin Guo School of Management, Qufu Normal University,Rizhao276826,China Abstract- Under the background of port supply chain strategic alliances,we clarify the current situation of Rizhao Port by SWOT analysis and find out the ST strategy for it, furthermore, we screen out the optimal cooperation partner for Rizhao Port under different alliance forms range from horizontal integration and vertical integration to the blended dynamic logistics alliance. Index Terms- Port Supply Chain; the SWOT Analysis; RiZhao Port; Strategical alliance I. INTRODUCTION ort supply chain, a main constituent of port logistics, includes the levels such as informationization, automation and networking, P and emphasizes the modern management of material transportation chain of each link and the extension of comprehensive services.Meanwhile, due to the connection between ports and suppliers and consumers all over the world through shippping companies and land forwarding agents, a port supply chain integrating many means1 of transportation and types of logistics is formed, and it becomes a relatively best main part and link for the coordinated management of supply chain. Therefore, we can start from the analysis of the SWOT of Rizhao port supply chain, and find out the cooperative partners suitable for the strategical alliance of Rizhao port supply chain among different types of alliance. II. THE SWOT ANALYSIS OF RIZHAO PORT SUPPLY CHAIN The SWOT analysis, a commonly used method of strategic competition, is based on analyzing the internal conditions of enterprises itself, and finds out the strengths, weaknesses and core competence. -

Land-Use Efficiency in Shandong (China)
sustainability Article Land-Use Efficiency in Shandong (China): Empirical Analysis Based on a Super-SBM Model Yayuan Pang and Xinjun Wang * Department of Environmental Science and Engineering, Fudan University, Shanghai 200433, China; [email protected] * Correspondence: [email protected] Received: 20 November 2020; Accepted: 14 December 2020; Published: 18 December 2020 Abstract: A reasonable evaluation of land-use efficiency is an important issue in land use and development. By using a super-SBM model, the construction and cultivated land-use efficiency of 17 cities in Shandong from 2006 to 2018 were estimated and the spatial-temporal variation was analyzed. The results showed that: (1) The land use efficiency levels were quite different, and low-efficiency cities impacted the overall development process. (2) The efficiency values of construction land generally fluctuated and rose, meaning that room remains for future efficiency improvements. Cultivated land generally showed a high utilization efficiency, but it fluctuated and decreased. (3) The construction land-use efficiency was highest in the midland region, especially in Laiwu city, followed by the eastern region and Qingdao city, and the western region. The spatial variation in cultivated land presented a trend of “high in the middle, low in the periphery,” centered on Jinan and Yantai city. (4) Pure technical efficiency was the main restriction driving inefficient utilization in the western region, while scale efficiency played that role in the east. Based on the findings, policy suggestions were proposed to improve the land-use efficiency in Shandong and promote urban sustainable development. Keywords: land use; efficiency level; super-SBM model; Shandong Province; construction land; cultivated land 1. -

Spatial and Temporal Variation of VIIRS Derived AOT
Spatial and temporal variation of the Visible Infrared Imaging Radiometer Suite (VIIRS) derived aerosol optical thickness over East China Fei Menga, b, Changyong Caoc, Xi Shaoa aDepartment of Astronomy, University of Maryland, College Park, MD, USA bDepartment of Civil Engineering, Shandong Jianzhu University, Jinan, PR China cNOAA/ NESDIS/STAR, College Park, MD, USA Abstract Results The spatial and temporal variations in regional aerosol optical thickness (AOT) were investigated over Annual and seasonal AOT variations for different cities Shandong province of China based upon one year’s Visible Infrared Imaging Radiometer Suite (VIIRS) Averaged over the measurements in 2013, the minimum (Min.), maximum (Max.), mean, standard deviation data. The regional forest background annual mean AOT was 0.467 with a standard deviation of 0.339, (Std.) and variance of AOT in 17 cities (Fig.1) are presented in Table 1. which was much higher than the background continental AOT level of 0.10. Higher AOT values for the In order to better understand the AOT values in different cities, percent days with AOT≤0.5, 1.0≥AOT>0.5 and study region were mainly found in the spring and summer, especially from May to August, while the AOT>1.0 respectively in each cities were calculated (Fig.2). (a) (b) (c) (d) lowest mean aerosol values were seen in November and December. Urban areas all have obviously higher mean AOT values than the rural areas resulting from intense anthropogenic sources. Given that the forest background AOT represents the natural background level, anthropogenic emissions and C secondary aerosol generation contribute approximately 0.352 to the aerosol loading in this region. -
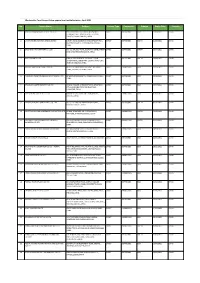
Apparel & Textile Factory List
Woolworths Food Group-Active apparel and textile factories- April 2021 No. Factory Name Address Factory Type Commodity Scheme Expiry Date Country 1 JIANGSU HONGMOFANG TEXTILE CO., LTD EAST CIFU ROAD, EAST RUISHENG AVENUE,, DIRECT SOFTGOODS BSCI 23/04/2021 CHINA ECONOMIC DEVELOPMENT ZONE, SHUYANG COUNTY,,SUQIAN,JIANGSU,,CHINA 2 LIANYUNGANG ZHAOWEN SHOES LIMITED 2 BEIHAI ROAD,ECONOMIC DEVELOPMENT AREA, DIRECT SOFTGOODS SMETA 11/05/2021 CHINA GUANNAN COUNTY,,LIANYUNGANG,JIANGSU,, CHINA 3 ZHUCHENG YIXIN GARMENT CO., LTD NO. 321 SHUNDU ROAD,,ECONOMIC DEVELOPMENT DIRECT SOFTGOODS SMETA 13/07/2021 CHINA ZONE,,ZHUCHENG,SHANDONG,,CHINA 4 HRX FASHION CO LTD 1000 METERS SOUTH OF GUOCANG TOWN DIRECT SOFTGOODS SMETA 25/08/2021 CHINA GOVERNMENT,,WENSHANG COUNTY, JINING CITY, JINING,SHANDONG,,CHINA 5 PUJIANG KINGSHOW CARPET CO LTD NO.75-1 ZHEN PU ROAD, PU JIANG, ZHE JIANG, DIRECT HARDGOODS BSCI 24/12/2021 CHINA CHINA,,PUJIANG,ZHEJIANG,,CHINA 6 YANGZHOU TENGYI SHOES MANUFACTURE CO., LTD. 13 WEST HONGCHENG RD,,,FANGXIANG,JIANGSU,, DIRECT SOFTGOODS BSCI 22/10/2021 CHINA CHINA 7 ZHAOYUAN CASTTE GARMENT CO LTD PANJIAJI VILLAGE, LINGLONG TOWN,,ZHAOYUAN DIRECT SOFTGOODS BSCI 06/07/2021 CHINA CITY, SHANDONG PROVINCE,ZHAOYUAN, SHANDONG,,CHINA 8 RUGAO HONGTAI TEXTILE CO LTD XINJIAN VILLAGE, JIANGAN TOWN,,RUGAO, DIRECT HARDGOODS BSCI 07/12/2021 CHINA JIANGSU,,CHINA 9 NANJING BIAOMEI HOMETEXTILES CO.,LTD NO.13 ERST WUCHU ROAD,HENGXI TOWN,,, DIRECT SOFTGOODS SMETA 04/11/2021 CHINA NANJING,JIANGSU,,CHINA 10 NANTONG YAOXING HOUSEWARE PRODUCTS CO.,LTD NO.999, TONGFUBEI RD., CHONGCHUAN, DIRECT HARDGOODS SMETA 24/09/2021 CHINA NANTONG,,NANTONG,JIANGSU,,CHINA 11 CHAOZHOU CHAOAN ZHENGYUN CERAMICS QIAO HU VILLAGE, CHAOAN, CHAOZHOU CITY, DIRECT HARDGOODS SMETA 19/05/2021 CHINA INDUSTRIAL CO LTD GUANGDONG, CHINA,,CHAOZHOU,GUANGDONG,, CHINA 12 YANTAI PACIFIC HOME FASHION FUSHAN MILL NO. -

Slip Op. 19-67 UNITED STATES COURT OF
Case 1:18-cv-00002-JCG Document 81 Filed 06/03/19 Page 1 of 30 Slip Op. 19-67 UNITED STATES COURT OF INTERNATIONAL TRADE LINYI CHENGEN IMPORT AND EXPORT CO., LTD., Plaintiff, and CELTIC CO., LTD. ET AL., Consolidated Plaintiffs, Before: Jennifer Choe-Groves, Judge v. Consol. Court No. 18-00002 UNITED STATES, Defendant, and COALITION FOR FAIR TRADE IN HARDWOOD PLYWOOD, Defendant-Intervenor. OPINION AND ORDER [Sustaining in part and remanding in part the U.S. Department of Commerce’s final determination in the antidumping duty investigation of certain hardwood plywood products from the People’s Republic of China.] Dated: June 3, 2019 Gregory S. Menegaz and Alexandra H. Salzman, deKieffer & Horgan, PLLC, of Washington, D.C., argued for Plaintiff Linyi Chengen Import and Export Co., Ltd., Consolidated Plaintiffs Celtic Co., Ltd., Jiaxing Gsun Import & Export Co., Ltd., Suqian Hopeway International Trade Co., Ltd., Anhui Hoda Wood Co., Ltd., Shanghai Futuwood Trading Co., Ltd., Linyi Evergreen Wood Co., Ltd., Xuzhou Jiangyang Wood Industries Co., Ltd., Xuzhou Timber International Trade Co. Ltd., Linyi Sanfortune Wood Co., Ltd., Linyi Mingzhu Wood Co., Ltd., Xuzhou Andefu Wood Co., Ltd., Suining Pengxiang Wood Co., Ltd., Xuzhou Shengping Import and Export Co., Ltd., Xuzhou Pinlin International Trade Co. Ltd., Linyi Glary Plywood Co., Ltd., Case 1:18-cv-00002-JCG Document 81 Filed 06/03/19 Page 2 of 30 Consol. Court No. 18-00002 Page 2 Linyi Linhai Wood Co., Ltd., Linyi Hengsheng Wood Industry Co., Ltd., Shandong Qishan International Trading -

Annual Report of Shandong Hi-Speed Co. Ltd. of 2020
Annual Report 2020 Company Code: 600350 Abbreviation of Company: Shangdong Hi-Speed Annual Report of Shandong Hi-Speed Co. Ltd. of 2020 1/265 Annual Report 2020 Notes I. The Board of Directors, Board of Supervisors, directors, supervisors and executives of the Company guarantee the truthfulness, accuracy and completeness without any false or misleading statements or material omissions herein, and shall bear joint and several legal liabilities. II. Absent directors Post of absent director Name of absent director Reason for absence Name of delegate Independent Director Fan Yuejin Because of work Wei Jian III. Shinewing Certified Public Accountants (Special Partnership) has issued an unqualified auditor's report for the Company. IV. Mr. Sai Zhiyi, the head of the Company, Mr. Lyu Sizhong, Chief Accountant who is in charge of accounting affairs, Mr. Zhou Liang, and Chen Fang (accountant in charge) from ShineWing declared to guarantee the truthfulness, accuracy and completeness of the annual report. V. With respect to the profit distribution plan or common reserves capitalizing plan during the reporting period reviewed by the Board of Directors After being audited by Shinewing Certified Public Accountants (Special Partnership), the net profit attributable to owners of the parent company in 2020 after consolidation is CNY 2,038,999,018.13, where: the net profit achieved by the parent company is CNY2,242,060,666.99. After withdrawing the statutory reserves of CNY224,206,066.70 at a ratio of 10% of the achieved net profit of the parent company, the retained earnings is 2,017,854,600.29 . The accumulated distributable profits of parent company in 2020 is CNY16,232,090,812.89. -
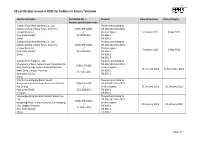
CE Certificates Issued in 2021 for Holders in China / Vietnam
CE certificates issued in 2021 for holders in China / Vietnam Certificate holder Certificate No. / Product Date of issuance Date of expiry Product specification code Xuzhou Yizun New Material Co., Ltd. Plywood according to Industrial Park, Hegou Town, Xinyi City, 0766-CPR-429/4 EN 636:2012+A1:2015 Jiangsu Province / Product types: 5 January 2021 9 May 2021 Post code 221439 2117181-009 EN 636-1 China EN 636-2 Xuzhou Yizun New Material Co., Ltd. Plywood according to Industrial Park, Hegou Town, Xinyi City, 0766-CPR-430/4 EN 636:2012+A1:2015 Jiangsu Province / Product types: 5 January 2021 9 May 2021 Post code 221439 2117181-010 EN 636-1 China EN 636-2 EN 636-3 Xuzhou Kinri Trade Co., Ltd. Plywood according to Shengyang Village, Sanpu Town, Tongshan Dis- EN 636:2012+A1:2015 0766-CPR-564 trict, Xuzhou High-tech Industrial Develop- Product types: / 11 January 2021 24 November 2021 ment Zone, Jiangsu Province EN 636-1 2121007-005 Post code 221112 EN 636-2 China Viet Nam Hai Duong Baifar Wood Plywood according to Nam Sach Industrial Zone, Nam Sach District, 0766-CPR-565 EN 636:2012+A1:2015 Hai Duong / Product types: 11 January 2021 10 January 2022 Post code 03000 2121008-001 EN 636-1 Vietnam EN 636-2 LianyungangChanta International Wood Cov Plywood according to Ltd. EN 636:2012+A1:2015 0766-CPR-485/2 Kangpeng Plaza, Lianyun District, Lianyungang Product types: / 26 January 2021 20 January 2022 City, Jiangsu Province EN 636-1 2119007-003 Post code 222000 EN 636-2 China EN 636-3 Page 1 of 7 CE certificates issued in 2021 for holders in China / Vietnam Certificate holder Certificate No. -

Evaluation on Sustainable Utilization of Water Resources in Shandong
2017 International Conference on Energy, Power and Environmental Engineering (ICEPEE 2017) ISBN: 978-1-60595-456-1 Evaluation on Sustainable Utilization of Water Resources in Shandong Province Based on Water Footprint Yu-heng YANG, Sheng-le CAO*, Chun-tong LIU and Yu-fei CHENG School of Civil Engineering, Shandong University, Jinan 250061, China Keywords. Water footprint, Water resources, Evaluation of sustainable development, Shandong province. Abstract. The paper adopts 17 prefecture-level cities in Shandong Province as geographical unit, selects the indicators concerning regional sustainable development capacity from the perspective of resource, environment, social economy and efficiency support, etc., introduces the concepts of water footprint and evaluates the sustainable development condition of water resource and overall sustainable development condition in these geographical units. It’s indicated that the overall sustainable development capacity of the 17 prefecture-level cities in Shandong Province is ranked in descending order as follows: Weihai, Yantai, Qingdao, Binzhou, Zibo, Dezhou, Jining, Rizhao, Jinan, Laiwu, Linyi, Zaozhuang, Weifang, Dongying, Liaocheng, Taian and Heze. The score of sustainable development capacity achieved by the former nine is larger than 0 and the cities are in a state of sustainable development; while the score of sustainable development capacity achieved by the latter eight is smaller than 0 and the cities are in a state of unsustainable development. On this basis, the geographical units are further divided into four types by means of optimal partition method, i.e., with strong sustainable development capacity, with comparatively strong sustainable development capacity, with comparatively weak sustainable development capacity and with weak sustainable development capacity, and the sustainable development characteristics of each type are analyzed. -

Download File
On the Periphery of a Great “Empire”: Secondary Formation of States and Their Material Basis in the Shandong Peninsula during the Late Bronze Age, ca. 1000-500 B.C.E Minna Wu Submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the Graduate School of Arts and Sciences COLUMIBIA UNIVERSITY 2013 @2013 Minna Wu All rights reserved ABSTRACT On the Periphery of a Great “Empire”: Secondary Formation of States and Their Material Basis in the Shandong Peninsula during the Late Bronze-Age, ca. 1000-500 B.C.E. Minna Wu The Shandong region has been of considerable interest to the study of ancient China due to its location in the eastern periphery of the central culture. For the Western Zhou state, Shandong was the “Far East” and it was a vast region of diverse landscape and complex cultural traditions during the Late Bronze-Age (1000-500 BCE). In this research, the developmental trajectories of three different types of secondary states are examined. The first type is the regional states established by the Zhou court; the second type is the indigenous Non-Zhou states with Dong Yi origins; the third type is the states that may have been formerly Shang polities and accepted Zhou rule after the Zhou conquest of Shang. On the one hand, this dissertation examines the dynamic social and cultural process in the eastern periphery in relation to the expansion and colonization of the Western Zhou state; on the other hand, it emphasizes the agency of the periphery during the formation of secondary states by examining how the polities in the periphery responded to the advances of the Western Zhou state and how local traditions impacted the composition of the local material assemblage which lay the foundation for the future prosperity of the regional culture. -

CHINA VANKE CO., LTD.* 萬科企業股份有限公司 (A Joint Stock Company Incorporated in the People’S Republic of China with Limited Liability) (Stock Code: 2202)
Hong Kong Exchanges and Clearing Limited and The Stock Exchange of Hong Kong Limited take no responsibility for the contents of this announcement, make no representation as to its accuracy or completeness and expressly disclaim any liability whatsoever for any loss howsoever arising from or in reliance upon the whole or any part of the contents of this announcement. CHINA VANKE CO., LTD.* 萬科企業股份有限公司 (A joint stock company incorporated in the People’s Republic of China with limited liability) (Stock Code: 2202) 2019 ANNUAL RESULTS ANNOUNCEMENT The board of directors (the “Board”) of China Vanke Co., Ltd.* (the “Company”) is pleased to announce the audited results of the Company and its subsidiaries for the year ended 31 December 2019. This announcement, containing the full text of the 2019 Annual Report of the Company, complies with the relevant requirements of the Rules Governing the Listing of Securities on The Stock Exchange of Hong Kong Limited in relation to information to accompany preliminary announcement of annual results. Printed version of the Company’s 2019 Annual Report will be delivered to the H-Share Holders of the Company and available for viewing on the websites of The Stock Exchange of Hong Kong Limited (www.hkexnews.hk) and of the Company (www.vanke.com) in April 2020. Both the Chinese and English versions of this results announcement are available on the websites of the Company (www.vanke.com) and The Stock Exchange of Hong Kong Limited (www.hkexnews.hk). In the event of any discrepancies in interpretations between the English version and Chinese version, the Chinese version shall prevail, except for the financial report prepared in accordance with International Financial Reporting Standards, of which the English version shall prevail.