OSME List V3.4 Passerines-2
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Morocco SD 2017 Trip Report
Morocco 9th - 18th March 2017 Desert Sparrow is surely one of the best looking and most sought after of all the sparrows Tour Leader: Lisle Gwynn All photos in this report were taken by Lisle Gwynn on this tour Species depicted in photographs are named in BOLD RED www.tropicalbirding.com +1-409-515-9110 [email protected] Introduction Morocco is a fascinating destination, and one that many world birders have neglected for too long. It is increasingly becoming a go-to country for European birders in Spring, and offers some of the most exciting birding in the Western Palearctic biogeographic region. Not only does it offer a chance to see Afro-European migration at its peak, but it also offers a plethora of exciting and special endemic and near-endemic species at its core. Add to this the fact that throughout the tour we have excellent accommodation and some of the best food available anywhere in the world (in my opinion), it all goes toward making Morocco a must-visit location for any birder branching out into the world. It is also currently by far the safest North African country to visit, with little crime and none of the problems that plague the rest of the region, and therefore presents a comfortable and safe opportunity to experience North Africa. This year’s tour followed our tried and tested route, starting in the manic city of Marrakesh at a serene hotel amongst the craziness, a quick departure to the idyllic Ourika Valley and the high snow-capped peaks of Oukameiden and the high Atlas Mountains, before descending to the stony desert around Boumalne Dades and the ochre-cast dunes of the Sahara at Erg Chebbi. -

500 Natural Sciences and Mathematics
500 500 Natural sciences and mathematics Natural sciences: sciences that deal with matter and energy, or with objects and processes observable in nature Class here interdisciplinary works on natural and applied sciences Class natural history in 508. Class scientific principles of a subject with the subject, plus notation 01 from Table 1, e.g., scientific principles of photography 770.1 For government policy on science, see 338.9; for applied sciences, see 600 See Manual at 231.7 vs. 213, 500, 576.8; also at 338.9 vs. 352.7, 500; also at 500 vs. 001 SUMMARY 500.2–.8 [Physical sciences, space sciences, groups of people] 501–509 Standard subdivisions and natural history 510 Mathematics 520 Astronomy and allied sciences 530 Physics 540 Chemistry and allied sciences 550 Earth sciences 560 Paleontology 570 Biology 580 Plants 590 Animals .2 Physical sciences For astronomy and allied sciences, see 520; for physics, see 530; for chemistry and allied sciences, see 540; for earth sciences, see 550 .5 Space sciences For astronomy, see 520; for earth sciences in other worlds, see 550. For space sciences aspects of a specific subject, see the subject, plus notation 091 from Table 1, e.g., chemical reactions in space 541.390919 See Manual at 520 vs. 500.5, 523.1, 530.1, 919.9 .8 Groups of people Add to base number 500.8 the numbers following —08 in notation 081–089 from Table 1, e.g., women in science 500.82 501 Philosophy and theory Class scientific method as a general research technique in 001.4; class scientific method applied in the natural sciences in 507.2 502 Miscellany 577 502 Dewey Decimal Classification 502 .8 Auxiliary techniques and procedures; apparatus, equipment, materials Including microscopy; microscopes; interdisciplinary works on microscopy Class stereology with compound microscopes, stereology with electron microscopes in 502; class interdisciplinary works on photomicrography in 778.3 For manufacture of microscopes, see 681. -

Cameroon 2003
Cameroon 2003 Ola Elleström Claes Engelbrecht Bengt Grandin Erling Jirle Nils Kjellén Jonas Nordin Bengt-Eric Sjölinder Sten Stemme Dan Zetterström Front cover: Mount Kupé Bushshrike, Telephorus kupeensis, by Dan Zetterström Cameroon map: Jonas Nordin INTRODUCTION AND PLANNING. By Erling Jirle FACTS ABOUT THE COUNTRY The population is about 11 millions. There are over 200 ethnic groups, in the southeast pygmies for example. In the north Moslems are in majority, and in the south Christians. Official languages are French and English. In most of the country French is the dominant language, English is spoken mainly in the southwest part of the country, in the former English colony. The flora consists of over 8000 known species. In the rainforest belt you can find 22 primate species (like Gorilla, Chimpanzee, Drill, Mandrill) and 22 antelopes. There are 7 National Parks and several large fauna reserves. In all 4,5 percent of the land area are reserves. CLIMATE The climate in Cameroon is complicated, since it comprises of several climate zones. All Cameroon is tropical. Annual mean temperature is 23-28 depending on altitude. In the North the rains are between June - September (400 mm), then Waza National Park usually becomes impassable. In the inner parts of Cameroon there are two ”rains”; May - June and Oct. - Nov. (1500 mm annually). The rainy season along the coast is around 8 months, roughly April - November (3800 mm). West of Mount Cameroon you find the third wettest spot on earth, with over 10 000 mm per year. Also the Western Highlands gets almost 10 meter rain between May - October. -

Comments on the Ornithology of Nigeria, Including Amendments to the National List
Robert J. Dowsett 154 Bull. B.O.C. 2015 135(2) Comments on the ornithology of Nigeria, including amendments to the national list by Robert J. Dowsett Received 16 December 2014 Summary.—This paper reviews the distribution of birds in Nigeria that were not treated in detail in the most recent national avifauna (Elgood et al. 1994). It clarifies certain range limits, and recommends the addition to the Nigerian list of four species (African Piculet Verreauxia africana, White-tailed Lark Mirafra albicauda, Western Black-headed Batis Batis erlangeri and Velvet-mantled Drongo Dicrurus modestus) and the deletion (in the absence of satisfactory documentation) of six others (Olive Ibis Bostrychia olivacea, Lesser Short-toed Lark Calandrella rufescens, Richard’s Pipit Anthus richardi, Little Grey Flycatcher Muscicapa epulata, Ussher’s Flycatcher M. ussheri and Rufous-winged Illadopsis Illadopsis rufescens). Recent research in West Africa has demonstrated the need to clarify the distributions of several bird species in Nigeria. I have re-examined much of the literature relating to the country, analysed the (largely unpublished) collection made by Boyd Alexander there in 1904–05 (in the Natural History Museum, Tring; NHMUK), and have reviewed the data available in the light of our own field work in Ghana (Dowsett-Lemaire & Dowsett 2014), Togo (Dowsett-Lemaire & Dowsett 2011a) and neighbouring Benin (Dowsett & Dowsett- Lemaire 2011, Dowsett-Lemaire & Dowsett 2009, 2010, 2011b). The northern or southern localities of species with limited ranges in Nigeria were not always detailed by Elgood et al. (1994), although such information is essential for understanding distribution patterns and future changes. For many Guineo-Congolian forest species their northern limit in West Africa lies on the escarpment of the Jos Plateau, especially Nindam Forest Reserve, Kagoro. -

The Birds (Aves) of Oromia, Ethiopia – an Annotated Checklist
European Journal of Taxonomy 306: 1–69 ISSN 2118-9773 https://doi.org/10.5852/ejt.2017.306 www.europeanjournaloftaxonomy.eu 2017 · Gedeon K. et al. This work is licensed under a Creative Commons Attribution 3.0 License. Monograph urn:lsid:zoobank.org:pub:A32EAE51-9051-458A-81DD-8EA921901CDC The birds (Aves) of Oromia, Ethiopia – an annotated checklist Kai GEDEON 1,*, Chemere ZEWDIE 2 & Till TÖPFER 3 1 Saxon Ornithologists’ Society, P.O. Box 1129, 09331 Hohenstein-Ernstthal, Germany. 2 Oromia Forest and Wildlife Enterprise, P.O. Box 1075, Debre Zeit, Ethiopia. 3 Zoological Research Museum Alexander Koenig, Centre for Taxonomy and Evolutionary Research, Adenauerallee 160, 53113 Bonn, Germany. * Corresponding author: [email protected] 2 Email: [email protected] 3 Email: [email protected] 1 urn:lsid:zoobank.org:author:F46B3F50-41E2-4629-9951-778F69A5BBA2 2 urn:lsid:zoobank.org:author:F59FEDB3-627A-4D52-A6CB-4F26846C0FC5 3 urn:lsid:zoobank.org:author:A87BE9B4-8FC6-4E11-8DB4-BDBB3CFBBEAA Abstract. Oromia is the largest National Regional State of Ethiopia. Here we present the first comprehensive checklist of its birds. A total of 804 bird species has been recorded, 601 of them confirmed (443) or assumed (158) to be breeding birds. At least 561 are all-year residents (and 31 more potentially so), at least 73 are Afrotropical migrants and visitors (and 44 more potentially so), and 184 are Palaearctic migrants and visitors (and eight more potentially so). Three species are endemic to Oromia, 18 to Ethiopia and 43 to the Horn of Africa. 170 Oromia bird species are biome restricted: 57 to the Afrotropical Highlands biome, 95 to the Somali-Masai biome, and 18 to the Sudan-Guinea Savanna biome. -

Birds Along Lehi's Trail
Journal of Book of Mormon Studies Volume 15 Number 2 Article 10 7-31-2006 Birds Along Lehi's Trail Stephen L. Carr Follow this and additional works at: https://scholarsarchive.byu.edu/jbms BYU ScholarsArchive Citation Carr, Stephen L. (2006) "Birds Along Lehi's Trail," Journal of Book of Mormon Studies: Vol. 15 : No. 2 , Article 10. Available at: https://scholarsarchive.byu.edu/jbms/vol15/iss2/10 This Feature Article is brought to you for free and open access by the Journals at BYU ScholarsArchive. It has been accepted for inclusion in Journal of Book of Mormon Studies by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. Title Birds Along Lehi’s Trail Author(s) Stephen L. Carr Reference Journal of Book of Mormon Studies 15/2 (2006): 84–93, 125–26. ISSN 1065-9366 (print), 2168-3158 (online) Abstract When Carr traveled to the Middle East, he observed the local birds. In this article, he suggests the possi- bility that the Book of Mormon prophet Lehi and his family relied on birds for food and for locating water. Carr discusses the various birds that Lehi’s family may have seen on their journey and the Mosaic law per- taining to those birds. Birds - ALOnG LEHI’S TRAIL stephen l. cARR 84 VOLUME 15, NUMBER 2, 2006 PHOTOGRAPHy By RICHARD wELLINGTOn he opportunity to observe The King James translators apparently ex- birds of the Middle East came to perienced difficulty in knowing exactly which me in September 2000 as a member Middle Eastern birds were meant in certain pas- Tof a small group of Latter-day Saints1 traveling in sages of the Hebrew Bible. -

Dieter Thomas Tietze Editor How They Arise, Modify and Vanish
Fascinating Life Sciences Dieter Thomas Tietze Editor Bird Species How They Arise, Modify and Vanish Fascinating Life Sciences This interdisciplinary series brings together the most essential and captivating topics in the life sciences. They range from the plant sciences to zoology, from the microbiome to macrobiome, and from basic biology to biotechnology. The series not only highlights fascinating research; it also discusses major challenges associated with the life sciences and related disciplines and outlines future research directions. Individual volumes provide in-depth information, are richly illustrated with photographs, illustrations, and maps, and feature suggestions for further reading or glossaries where appropriate. Interested researchers in all areas of the life sciences, as well as biology enthusiasts, will find the series’ interdisciplinary focus and highly readable volumes especially appealing. More information about this series at http://www.springer.com/series/15408 Dieter Thomas Tietze Editor Bird Species How They Arise, Modify and Vanish Editor Dieter Thomas Tietze Natural History Museum Basel Basel, Switzerland ISSN 2509-6745 ISSN 2509-6753 (electronic) Fascinating Life Sciences ISBN 978-3-319-91688-0 ISBN 978-3-319-91689-7 (eBook) https://doi.org/10.1007/978-3-319-91689-7 Library of Congress Control Number: 2018948152 © The Editor(s) (if applicable) and The Author(s) 2018. This book is an open access publication. Open Access This book is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made. -
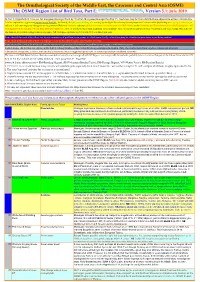
ORL 5.1 Hypothetical Spp Final Draft01a.Xlsx
The Ornithological Society of the Middle East, the Caucasus and Central Asia (OSME) The OSME Region List of Bird Taxa, Part E: , Version 5.1: July 2019 In Part E, Hypothetical Taxa, we list non-passerines (prefixed by 'N') first, then passerines (prefixed by 'P'). Such taxa may be from distributions adjacent to or have extended to A fuller explanation is given in Explanation of the ORL, but briefly, Bright green shading of a row (eg Syrian Ostrich) indicates former presence of a taxon in the OSME Region. Light gold shading in column A indicates sequence change from the previous ORL issue. Red font indicates added information since the previous ORL version or the Conservation Threat Status (Critically Endangered = CE, Endangered = E, Vulnerable = V and Data Deficient = DD only). Not all synonyms have been examined. Serial numbers (SN) are merely an administrative convenience and may change. Please do not cite them in any formal correspondence or papers. NB: Compass cardinals (eg N = north, SE = southeast) are used. Rows shaded thus and with yellow text denote summaries of problem taxon groups in which some closely-related taxa may be of indeterminate status or are being studied. Rows shaded thus and with yellow text indicate recent or data-driven major conservation concerns. Rows shaded thus and with white text contain additional explanatory information on problem taxon groups as and when necessary. English names shaded thus are species on BirdLife Tracking Database, http://seabirdtracking.org/mapper/index.php. Only a few individuals from very few colonies are involved. A broad dark orange line, as below, indicates the last taxon in a new or suggested species split, or where sspp are best considered separately. -

Erling Jirle Och Markus Lagerqvist, Tk, April 2019 1 VETENSKAPLIGT NAMN SVENSKT NAMN ENGELS
STATUS i Sverige VETENSKAPLIGT NAMN SVENSKT NAMN ENGELSKT NAMN Noter (Scientific name) (Swedish name) (English name) (Notes) Ordning STRUTHIONIFORMES STRUTSFÅGLAR Familj Struthionidae Strutsar Ostriches Struthio camelus struts Common Ostrich Ordning ANSERIFORMES ANDFÅGLAR Familj Anatidae Änder Ducks, Geese and Swans Dendrocygna bicolor brun visseland Fulvous Whistling Duck Dendrocygna javanica orientvisseland Lesser Whistling Duck M Branta bernicla prutgås Brant Goose R Branta ruficollis rödhalsad gås Red-breasted Goose B Branta canadensis kanadagås Canada Goose B Branta leucopsis vitkindad gås Barnacle Goose (E) Branta hutchinsii dvärgkanadagås Cackling Goose (D) R Anser indicus stripgås Bar-headed Goose (D) 14 Anser rossii dvärgsnögås Ross's Goose (E) R Anser caerulescens snögås Snow Goose B Anser anser grågås Greylag Goose B Anser fabalis sädgås Bean Goose M Anser brachyrhynchus spetsbergsgås Pink-footed Goose M Anser albifrons bläsgås Greater White-fronted Goose B Anser erythropus fjällgås Lesser White-fronted Goose (E) R Cygnus atratus svart svan Black Swan Intr. B Cygnus olor knölsvan Mute Swan M Cygnus columbianus mindre sångsvan Tundra Swan B Cygnus cygnus sångsvan Whooper Swan Plectropterus gambensis sporrgås Spur-winged Goose Sarkidiornis melanotos knöland Comb Duck R Alopochen aegyptiaca nilgås Egyptian Goose B Tadorna tadorna gravand Common Shelduck R Tadorna ferruginea rostand Ruddy Shelduck (E) 83 Aix sponsa brudand Wood Duck (E) b Aix galericulata mandarinand Mandarin Duck Intr. Nettapus coromandelianus bomullsdvärgand Cotton -

Biodiversity Profile of Afghanistan
NEPA Biodiversity Profile of Afghanistan An Output of the National Capacity Needs Self-Assessment for Global Environment Management (NCSA) for Afghanistan June 2008 United Nations Environment Programme Post-Conflict and Disaster Management Branch First published in Kabul in 2008 by the United Nations Environment Programme. Copyright © 2008, United Nations Environment Programme. This publication may be reproduced in whole or in part and in any form for educational or non-profit purposes without special permission from the copyright holder, provided acknowledgement of the source is made. UNEP would appreciate receiving a copy of any publication that uses this publication as a source. No use of this publication may be made for resale or for any other commercial purpose whatsoever without prior permission in writing from the United Nations Environment Programme. United Nations Environment Programme Darulaman Kabul, Afghanistan Tel: +93 (0)799 382 571 E-mail: [email protected] Web: http://www.unep.org DISCLAIMER The contents of this volume do not necessarily reflect the views of UNEP, or contributory organizations. The designations employed and the presentations do not imply the expressions of any opinion whatsoever on the part of UNEP or contributory organizations concerning the legal status of any country, territory, city or area or its authority, or concerning the delimitation of its frontiers or boundaries. Unless otherwise credited, all the photos in this publication have been taken by the UNEP staff. Design and Layout: Rachel Dolores -

Buckbird Journeys
BUCKBIRD JOURNEYS YEMEN and SOCOTRA Tuesday 8 – Sunday 18 November 2007 Participants Louise Augustine (LA), Hugh Buck (HB), David Daniels (DD), David Hoddinott (DH), Pearl Jordan (PJ), Werner Suter (WS), David Bradford (DB – Socotra only) This trip was designed to try for all the Southwest Arabia and Socotra endemics in a relatively brief time scale. It also represented an opportunity for regional listers to add an enticing selection of “African” species only entering Asia in Southwest Arabia and a selection of “Asian” species found in Africa rarely outside of Socotra. That the trip was successful on all counts is a tribute to Yousuf Mohageb (YM) of Arabian Eco- Tours in Sana’a who designed, with HB, the itinerary, accompanied us throughout Yemen and knew all the best birding spots, Ali his redoubtable co-driver and Ahmed Saeid Suliman (AS) on Socotra whose knowledge and love of all things on his island home is second to none. Our multinational group (three Americans, two Brits, a South African and a Swiss) kept to the pace admirably, provided exceptional spotting skills and, in WS, some wonderful photographic documentation and memories. Day by Day Thursday 8 November HB, DD and DH meet up at Dubai International Airport for the short Emirates Airlines flight to Sana’a where they are met by YM, Ali in traditional Yemeni costume and LA and PJ who have arrived the previous evening. Under the high bright sun, which will be the norm for the next 10 days, we traverse Sana’a’s sprawling outskirts to the Funduk Arabia Felix, an interesting hotel made up a several old traditional houses right at the edge of the historic and UNESCO rated old town. -

Songbird Remix Sparrows of the World
Avian Models for 3D Applications Characters and Texture Mapping by Ken Gilliland 1 Songbird ReMix Sparrows of the World Contents Manual Introduction 3 Overview and Use 3 Creating a Songbird ReMix Bird with Poser or DAZ Studio 4 One Folder to Rule Them All 4 Physical-based Rendering 5 Posing & Shaping Considerations 5 Where to Find Your Birds and Poses 6 Field Guide List of Species 7 Old World Sparrows Spanish Sparrow 8 Italian Sparrow 10 Eurasian Tree Sparrow 12 Dead Sea Sparrow 14 Arabian Golden Sparrow 16 Russet Sparrow 17 Cape Sparrow 19 Great Sparrow 21 Chestnut Sparrow 23 New World Sparrows American Tree Sparrow 25 Harris's Sparrow 28 Fox Sparrow 30 Golden-crowned Sparrow 32 Lark Sparrow 35 Lincoln's Sparrow 37 Rufous-crowned Sparrow 39 Savannah Sparrow 43 Rufous-winged Sparrow 47 Resources, Credits and Thanks 49 Copyrighted 2013-20 by Ken Gilliland www.songbirdremix.com Opinions expressed on this booklet are solely that of the author, Ken Gilliland, and may or may not reflect the opinions of the publisher. 2 Songbird ReMix Sparrows of the World Introduction Sparrows are probably the most familiar of all wild birds. Throughout history sparrows have been considered the harbinger of good or bad luck. They are referred to in many works of ancient literature and religious texts around the world. The ancient Egyptians used the sparrow symbol in their hieroglyphs to express evil tidings, the ancient Greeks associated it with Aphrodite, the goddess of love as a lustful messenger, and Jesus used sparrows as an example of divine providence in the Gospel of Matthew.