Motor Systems Spinal-Cortex
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Revised Computational Neuroanatomy for Motor Control
This is the author’s final version; this article has been accepted for publication in the Journal of Cognitive Neuroscience 1 A Revised Computational Neuroanatomy for Motor Control 2 Shlomi Haar1, Opher Donchin2,3 3 1. Department of BioEngineering, Imperial College London, UK 4 2. Department of Biomedical Engineering, Ben-Gurion University of the Negev, Israel 5 3. Zlotowski Center for Neuroscience, Ben-Gurion University of the Negev, Israel 6 7 Corresponding author: Shlomi Haar ([email protected]) 8 Imperial College London, London, SW7 2AZ, UK 9 10 Acknowledgements: We would like to thank Ilan Dinstein, Liad Mudrik, Daniel Glaser, Alex Gail, and 11 Reza Shadmehr for helpful discussions about the manuscript. Shlomi Haar is supported by the Royal 12 Society – Kohn International Fellowship (NF170650). Work on this review was partially supported by 13 DFG grant TI-239/16-1. 14 15 Abstract 16 We discuss a new framework for understanding the structure of motor control. Our approach 17 integrates existing models of motor control with the reality of hierarchical cortical processing and the 18 parallel segregated loops that characterize cortical-subcortical connections. We also incorporate the recent 19 claim that cortex functions via predictive representation and optimal information utilization. Our 20 framework assumes each cortical area engaged in motor control generates a predictive model of a different 21 aspect of motor behavior. In maintaining these predictive models, each area interacts with a different part 22 of the cerebellum and basal ganglia. These subcortical areas are thus engaged in domain appropriate 23 system identification and optimization. This refocuses the question of division of function among different 24 cortical areas. -
Corticospinal Fibers
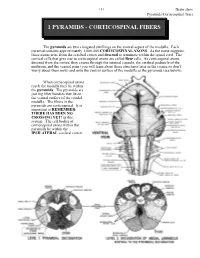
151 Brain stem Pyramids/Corticospinal Tract 1 PYRAMIDS - CORTICOSPINAL FIBERS The pyramids are two elongated swellings on the ventral aspect of the medulla. Each pyramid contains approximately 1,000,000 CORTICOSPINAL AXONS. As the name suggests, these axons arise from the cerebral cortex and descend to terminate within the spinal cord. The cortical cells that give rise to corticospinal axons are called Betz cells. As corticospinal axons descend from the cortex, they course through the internal capsule, the cerebral peduncle of the midbrain, and the ventral pons (you will learn about these structures later in the course so don’t worry about them now) and onto the ventral surface of the medulla as the pyramids (see below). When corticospinal axons reach the medulla they lie within the pyramids. The pyramids are just big fiber bundles that lie on the ventral surface of the caudal medulla. The fibers in the pyramids are corticospinal. It is important to REMEMBER: THERE HAS BEEN NO CROSSING YET! in this system. The cell bodies of corticospinal axons within the pyramids lie within the IPSILATERAL cerebral cortex. Brain stem 152 Pyramids/Corticospinal Tract At the most caudal pole of the pyramids the corticospinal axons cross over the midline and now continue their descent on the contralateral (to the cell of origin) side. This crossover point is called the PYRAMIDAL DECUSSATION. The crossing fibers enter the lateral funiculus of the spinal cord where they are called the LATERAL CORTICOSPINAL TRACT (“corticospinal” is not good enough, you have to call them lateral corticospinal; LCST - remember this one??). LCST axons exit the tract to terminate upon neurons in the spinal cord gray matter along its entire length. -

Motor Control: a Sense of Movement
RESEARCH HIGHLIGHTS MOTOR CONTROL A sense of movement Neuroscience textbooks tell us that with a shorter latency after S1 stimu- whisker protraction, and that M1retract the motor cortex controls move- lation than after M1 stimulation. only induces whisker retraction ment. But now, Carl Petersen and Thus, S1 and M1 are both involved in indirectly, through S1 activation. colleagues show that sensory cortex whisker motor and sensory process- By mapping the neural pathways may have an equally important role ing. The M1 area that induced C2 involved in C2 whisker retraction in motor control. retraction was termed M1retract; a and protraction, the authors found The authors showed that a single, more medial M1 region that induced that M1C2 and S1C2 have reciprocal brief deflection of the C2 whisker C2 protraction under microstimula- connections and project to adjacent induced activity in the correspond- tion was termed M1protract. regions in subcortical areas. In the ing barrel column of the primary The authors next investigated the brain stem, this includes the reticular somatosensory cortex (S1), followed functional relevance of this finding. formation as a projection area of M1, by a response in a small area in the By attaching metal particles to the C2 and the spinal trigeminal nuclei as primary motor cortex (M1). Direct whisker and applying a pulsed mag- a projection area of S1. Both areas microstimulation or optogenetic netic field, the authors could evoke project to the facial nucleus, which stimulation of either area induced C2 whisker deflections. The whisker contains whisker motor neurons. a brief retraction of the C2 whisker, retracted in response to this stimulus, Indeed, direct electrical stimula- and this response was abolished when tion of the reticular formation and S1 was inactivated — but not spinal trigeminal nuclei induced when M1 was inactivated — with whisker protraction and retraction, tetrodoxin (TTX). -

Motor Control in the Brain
Motor Control in the Brain Travis DeWolf School of Computer Science University of Waterloo Waterloo, ON Canada December 19, 2008 Abstract There has been much progress in the development of a model of motor control in the brain in the last decade; from the improved method for mathematically extracting the predicted movement direction from a population of neurons to the application of optimal control theory to motor control models, much work has been done to further our understanding of this area. In this paper recent literature is reviewed and the direction of future research is examined. 1 Introduction In the last decade there has been a push from investigating the coordinate system used in the brain, which has been the focus of research since the 1970s, back toward using reductionist experiment techniques and developing more encompassing models of motor control in the brain[28]. Optimal control theory applied to motor control in the brain has also recently started being explored as a framework for models. This has lead to research into the underlying biology of the implementations of internal models of movement and reinforcement learning reward systems in the brain[32]. In addition to this, with the discovery of motor primitives[24], which are often referred to as the `building blocks of movement', many of the ideas about neural activity carried out in the motor cortex need be reworked. The analysis techniques employed by researchers have been improved and of course the computing power available has increased enormously, and these developments have in turn opened the door to the need for further mathematical technique developments for accurate movement analysis and prediction. -

Sensory Change Following Motor Learning
A. M. Green, C. E. Chapman, J. F. Kalaska and F. Lepore (Eds.) Progress in Brain Research, Vol. 191 ISSN: 0079-6123 Copyright Ó 2011 Elsevier B.V. All rights reserved. CHAPTER 2 Sensory change following motor learning { k { { Andrew A. G. Mattar , Sazzad M. Nasir , Mohammad Darainy , and { } David J. Ostry , ,* { Department of Psychology, McGill University, Montréal, Québec, Canada { Shahed University, Tehran, Iran } Haskins Laboratories, New Haven, Connecticut, USA k The Roxelyn and Richard Pepper Department of Communication Sciences and Disorders, Northwestern University, Evanston, Illinois, USA Abstract: Here we describe two studies linking perceptual change with motor learning. In the first, we document persistent changes in somatosensory perception that occur following force field learning. Subjects learned to control a robotic device that applied forces to the hand during arm movements. This led to a change in the sensed position of the limb that lasted at least 24 h. Control experiments revealed that the sensory change depended on motor learning. In the second study, we describe changes in the perception of speech sounds that occur following speech motor learning. Subjects adapted control of speech movements to compensate for loads applied to the jaw by a robot. Perception of speech sounds was measured before and after motor learning. Adapted subjects showed a consistent shift in perception. In contrast, no consistent shift was seen in control subjects and subjects that did not adapt to the load. These studies suggest that motor learning changes both sensory and motor function. Keywords: motor learning; sensory plasticity; arm movements; proprioception; speech motor control; auditory perception. Introduction the human motor system and, likewise, to skill acquisition in the adult nervous system. -

Motor Control Paper“ for Our Common Script - IPNFA - Nicola Fischer, Carsten Schaefer „Well-Fed-Version“
suggestion for a „Motor Control Paper“ for our common script - IPNFA - Nicola Fischer, Carsten Schaefer „well-fed-version“ _______________________________________ Motor Control _______________________________________ 1. Definition and Contributions of Motor Control 2. Postural control 3. Activities 1. Definitions and Contributions Ø Definition of Motor Control “Motor control is defined as the ability to regulate or direct the mechanisms essential to movement.” (Shumway-Cook & Woollacott 2011, p.4; Horak et al 1997) Relevant questions are: - “How does the central nervous system (CNS) organize the many individual muscles and joints into coordinated functional movements? (Bernstein 1967; Shumway-Cook & Woollacott 2011; Magill 2003; Schmidt & Lee 2011) - How is sensory information from the environment and the body used to select and control movement? - What is the best way to study movement, and how can movement problems be quantified in patients with motor control problems?” (Shumway-Cook & Woollacott 2011, p.4) “Movement emerges from interactions between the individual, the task and the environment.” (Shumway-Cook & Woollacott 2011, p.5) figure 1: adapted from Shumway-Cook (Shumway-Cook & Woollacott 2011) Ø Theories of Motor Control § Reflex Theory e.g. Sir. Ch. Sherrington (Sherrington 1947) § Hierarchical Theory e.g. Rudolf Magnus, Arnold Gesell § Motor Programming Theory e.g. Karl Lashley (Fitch & Martins 2014) Nicolai Bernstein … as a base for the Systems Theory (Bernstein 1967) § Systems Theory / Dynamic Action Theory / Dynamic Pattern Theory - self organizing systems e.g. J.A. Scott Kelso, Viktor Jirsa (Jirsa & Kelso 2004) § Ecological Theory e.g. James Gibson (Gibson 1983) Ø Sensory Contributions to Motor Control For the closed-loop control system, sensory (or afferent) information is necessary to regulate our suggestion for a „Motor Control Paper“ for our common script - IPNFA - Nicola Fischer, Carsten Schaefer „well-fed-version“ movements (Adams 1971; Schmidt & Lee 2011). -

Visualization of the Pyramidal Decussation Utilizing Diffusion
OPEN ACCESS RESEARCH Visualization of the pyramidal decussation utilizing diffusion tensor imaging: a feasibility study utilizing generalized q-sampling imaging Erik H Middlebrooks1,*, Jeffrey A Bennett1, Sharatchandra Bidari1, and Alissa Old Crow1 1Department of Radiology, University of Florida College of Medicine, Gainesville, Florida, USA *corresponding author ([email protected]) Abstract Background: The delineating point of the end of the brainstem and beginning of the spinal cord is known as the cervicomedullary junction (CMJ). This point is defined by the decussation of the pyramidal tracts. Abnormal configuration and location of the CMJ have both been implicated in disease processes such as Chiari malformation. Unfortunately, the CMJ is not directly visualized on contemporary imaging techniques. Diffusion tensor imaging (DTI) has given us the ability to directly visualize white matter tracts, but suffers from difficulties with visualizing crossing fibers. Many advanced techniques for visualizing crossing fibers utilize substantially long imaging times or non-clinical magnet strengths making clinical applicability limited. Objective: This study inves- tigates the use of generalized q-sampling imaging with diffusion decomposition on standard DTI acquisitions at 3 Tesla to demonstrate the pyramidal decussation. Methods: Three differing DTI protocols were analyzed in vivo with scan times of 17 minutes, 10 minutes, and 5.5 minutes. Data was processed with standard DTI post-processing, as well as generalized q-sampling imaging with diffusion decomposition. Results: The results of the study show that the pyramidal decussation can be reliably visualized using generalized q-sampling imaging and diffusion decomposition with scan times as low at 10 minutes. Conclusion: Utilizing generalized q-sampling post-processing, the pyramidal decussation can be reliably visualized using clinically feasible DTI sequences with scan times as low as 10 minutes. -

Auditory and Vestibular Systems Objective • to Learn the Functional
Auditory and Vestibular Systems Objective • To learn the functional organization of the auditory and vestibular systems • To understand how one can use changes in auditory function following injury to localize the site of a lesion • To begin to learn the vestibular pathways, as a prelude to studying motor pathways controlling balance in a later lab. Ch 7 Key Figs: 7-1; 7-2; 7-4; 7-5 Clinical Case #2 Hearing loss and dizziness; CC4-1 Self evaluation • Be able to identify all structures listed in key terms and describe briefly their principal functions • Use neuroanatomy on the web to test your understanding ************************************************************************************** List of media F-5 Vestibular efferent connections The first order neurons of the vestibular system are bipolar cells whose cell bodies are located in the vestibular ganglion in the internal ear (NTA Fig. 7-3). The distal processes of these cells contact the receptor hair cells located within the ampulae of the semicircular canals and the utricle and saccule. The central processes of the bipolar cells constitute the vestibular portion of the vestibulocochlear (VIIIth cranial) nerve. Most of these primary vestibular afferents enter the ipsilateral brain stem inferior to the inferior cerebellar peduncle to terminate in the vestibular nuclear complex, which is located in the medulla and caudal pons. The vestibular nuclear complex (NTA Figs, 7-2, 7-3), which lies in the floor of the fourth ventricle, contains four nuclei: 1) the superior vestibular nucleus; 2) the inferior vestibular nucleus; 3) the lateral vestibular nucleus; and 4) the medial vestibular nucleus. Vestibular nuclei give rise to secondary fibers that project to the cerebellum, certain motor cranial nerve nuclei, the reticular formation, all spinal levels, and the thalamus. -

The Perception for Action Control Theory (PACT): a Perceptuo-Motor Theory of Speech Perception Jean-Luc Schwartz, Anahita Basirat, Lucie Ménard, Marc Sato
The Perception for Action Control Theory (PACT): a perceptuo-motor theory of speech perception Jean-Luc Schwartz, Anahita Basirat, Lucie Ménard, Marc Sato To cite this version: Jean-Luc Schwartz, Anahita Basirat, Lucie Ménard, Marc Sato. The Perception for Action Control Theory (PACT): a perceptuo-motor theory of speech perception. Journal of Neurolinguistics, Elsevier, 2012, 25 (5), pp.336-354. 10.1016/j.jneuroling.2009.12.004. hal-00442367 HAL Id: hal-00442367 https://hal.archives-ouvertes.fr/hal-00442367 Submitted on 21 Dec 2009 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. The Perception for Action Control Theory (PACT): a perceptuo-motor theory of speech perception Jean-Luc Schwartz (1), Anahita Basirat (1), Lucie Ménard (2), Marc Sato (1) (1) GIPSA-Lab, Speech and Cognition Department (ICP), UMR 5216 CNRS – Grenoble University, France (2) Laboratoire de Phonétique, UQAM / CRLMB, Montreal, Canada Abstract It is an old-standing debate in the field of speech communication to determine whether speech perception involves auditory or multisensory representations and processing, independently on any procedural knowledge about the production of speech units or on the contrary if it is based on a recoding of the sensory input in terms of articulatory gestures, as posited in the Motor Theory of Speech Perception. -

Proprioception in Motor Learning: Lessons from a Deafferented Subject
Exp Brain Res DOI 10.1007/s00221-015-4315-8 RESEARCH ARTICLE Proprioception in motor learning: lessons from a deafferented subject N. Yousif1 · J. Cole2 · J. Rothwell3 · J. Diedrichsen4 Received: 11 July 2014 / Accepted: 7 May 2015 © Springer-Verlag Berlin Heidelberg 2015 Abstract Proprioceptive information arises from a vari- movement in the perturbed direction. This suggests that ety of channels, including muscle, tendon, and skin affer- this form of learning may depend on static position sense at ents. It tells us where our static limbs are in space and the end of the movement. Our results indicate that dynamic how they are moving. It remains unclear however, how and static proprioception play dissociable roles in motor these proprioceptive modes contribute to motor learning. learning. Here, we studied a subject (IW) who has lost large myeli- nated fibres below the neck and found that he was strongly Keywords Proprioception · Motor control · impaired in sensing the static position of his upper limbs, Deafferentation · Force field learning when passively moved to an unseen location. When making reaching movements however, his ability to discriminate in which direction the trajectory had been diverted was unim- Introduction paired. This dissociation allowed us to test the involve- ment of static and dynamic proprioception in motor learn- Proprioception is a collective term that refers to non-visual ing. We found that IW showed a preserved ability to adapt input that tells us where our body is in space. Propriocep- to force fields when visual feedback was present. He was tion has an important function in normal motor control even sensitive to the exact form of the force perturbation, and motor learning. -

Proprioception and Motor Control in Parkinson's Disease
Journal of Motor Behavior, Vol. 41, No. 6, 2009 Copyright C 2009 Heldref Publications Proprioception and Motor Control in Parkinson’s Disease Jurgen¨ Konczak1,6, Daniel M. Corcos2,FayHorak3, Howard Poizner4, Mark Shapiro5,PaulTuite6, Jens Volkmann7, Matthias Maschke8 1School of Kinesiology, University of Minnesota, Minneapolis. 2Department of Kinesiology and Nutrition, University of Illinois at Chicago. 3Department of Science and Engineering, Oregon Health and Science University, Portland. 4Institute for Neural Computation, University of California–San Diego. 5Department of Physical Medicine and Rehabilitation, Northwestern University, Chicago, Illinois. 6Department of Neurology, University of Minnesota, Minneapolis. 7Department of Neurology, Universitat¨ Kiel, Germany. 8Department of Neurology, Bruderkrankenhaus,¨ Trier, Germany. ABSTRACT. Parkinson’s disease (PD) is a neurodegenerative dis- cles, tendons, and joint capsules. These receptors provide order that leads to a progressive decline in motor function. Growing information about muscle length, contractile speed, muscle evidence indicates that PD patients also experience an array of tension, and joint position. Collectively, this latter informa- sensory problems that negatively impact motor function. This is es- pecially true for proprioceptive deficits, which profoundly degrade tion is also referred to as proprioception or muscle sense. motor performance. This review specifically address the relation According to the classical definition by Goldscheider (1898) between proprioception and motor impairments in PD. It is struc- the four properties of the muscle sense are (a) passive mo- tured around 4 themes: (a) It examines whether the sensitivity of tion sense, (b) active motion sense, (c) limb position sense, kinaesthetic perception, which is based on proprioceptive inputs, is and (d) the sense of heaviness. Alternatively, some use the actually altered in PD. -

Real-Time Motor Control Using Recurrent Neural Networks
Real-Time Motor Control using Recurrent Neural Networks Dongsung Huh, and Emanuel Todorov Abstract—In this paper, we introduce a novel neural network single task only. On the other hand, most motor areas, architecture for motor control. Our general framework employs a recurrent neural network (RNN) to govern a dynamical system (body) in a closed loop fashion. This hybrid system is trained to behave as an interpreter that translates high-level motor plan into desired movement. Our method uses a variant of optimal control theory applicable to neural networks. When applied to biology, our method yields recurrent neural networks which are analogous to circuits in primary motor cortex (M1) or central pattern generators (CPGs) in the spinal cord. Fig 1. Body is a dynamical system with state 풙. Controller receives time-varying command (task-information) 풌 and controls body accordingly. The combined body-controller system can be seen as I. INTRODUCTION performing a mapping from command sequence 풌 to body-state In recent years, there has been much progress in the study sequence 풙 , i.e. movement. of biological motor control, both experimentally and including the primary motor cortex, subserve multiple theoretically. Numerous neural recording experiments have motor-tasks under changing task environment, where the task yielded insight into the neural substrate of motor control. information (or command) is given from the decision making Optimality principles of sensory motor functions have been (or motor-planning) centers of the brain. Therefore, it is successful in explaining behavior [1,2]. However, there is a appropriate to model the motor area as performing a general lack of understanding of how the two are related.