Ecophysiology ΟΙ Germination Ίn the Aromatic Plants Thyme, Savory And
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Amendment of Solicitation/Modification of Contract 1 9
1. CONTRACT ID CODE PAGE OF PAGES AMENDMENT OF SOLICITATION/MODIFICATION OF CONTRACT 1 9 2. AMENDMENT/MODIFICATION NO. 3. EFFECTIVE DATE 4. REQUISITION/PURCHASE REQ. NO. 5. PROJECT NO. (If applicable) 0003 See Block 14 6. ISSUED BY CODE SPE600 7. ADMINISTERED BY (If other than Item 6) CODE DLA ENERGY 8725 JOHN J KINGMAN RD STE 4950 FT BELVOIR VA 22060-6222 USA 8. NAME AND ADDRESS OF CONTRACTOR (No., street, county, State and ZIP Code) (X) 9A. AMENDMENT OF SOLICITATION NO. SPE60016R0208 X 9B. DATED (SEE ITEM 11) 2015 DEC 02 10A. MODIFICATION OF CONTRACT/ORDER NO. 10B. DATED (SEE ITEM 13) CODE FACILITY CODE 11. THIS ITEM ONLY APPLIES TO AMENDMENTS OF SOLICITATIONS X The above numbered solicitation is amended as set forth in Item 14. The hour and date specified for receipt of Offers is extended, is not extended. Offers must acknowledge receipt of this amendment prior to the hour and date specified in the solicitation or as amended, by one of the following methods: (a) By completing Items 8 and 15, and returning 1 copies of the amendment; (b) By acknowledging receipt of this amendment on each copy of the offer submitted; or (c) By separate letter or telegram which includes a reference to the solicitation and amendment numbers. FAILURE OF YOUR ACKNOWLEDGMENT TO BE RECEIVED AT THE PLACE DESIGNATED FOR THE RECEIPT OF OFFERS PRIOR TO THE HOUR AND DATE SPECIFIED MAY RESULT IN REJECTION OF YOUR OFFER. If by virtue of this amendment you desire to change an offer already submitted, such change may be made by telegram or letter, provided each telegram or letter makes reference to the solicitation and this amendment, and is received prior to the opening hour and date specified. -

Crete Bird Report 2000 and 2001
CRETE BIRD REPORT 2000-2001 WEATHER: 2000: Sunny and overcast in the first week in April with a hot southern wind in the west on 5th, then bright anda cold wind for the rest of April until 18th when there was rain and cold winds for 3 days. It was clear and settled from then onwards until the first week in May when there were strong winds and storms and large numbers of migrants on the south coast were reported. From the 9th May the weather was more settled. Strong winds reported on Ist June, which brought in Gulls including Audouin’s at Elounda. In the Autimn, late September and early October, there was some bad weather with gale force winds, driving rain and very little sunshine. Passage of birds was noted on 7th and 8th October. The rest of the month was settled with occasional brief showers. 2001: April began cold and wet with temperatures of 13° recorded in Plakias, on 3rd, and cold and wet, bright intervals and then heavy weather, which brought in migrants. Weather still cool and unsettled until 10th, but good birding resulted. Late April was warmer and humid with thunderstorms and Sahara sand, then sunny but persistent fresh blustery N W winds. During the last week of April the weather settled at last with temperatures of 27° and continued hot with strong northerly winds, then settling by 8th May when it became hot with cloudless skies. The Autumn was hot in September and October. The weather broke in early November with the winter rains arriving. -

List of Bank Branches (By Hebic) 31/03/2021
LIST OF BANK BRANCHES (BY HEBIC) 31/03/2021 BANK OF GREECE HEBIC BRANCH NAME AREA ADDRESS TELEPHONE NUMBER / FAX NUMBER 0100001 HEAD OFFICE SECRETARIAT ATHENS CENTRE 21, El. Venizelou Ave., 102 50 ATHENS SECTION tel.: 2103202049, fax: 2103226371 0100002 HEAD OFFICE TENDER AND ATHENS CENTRE 21, El. Venizelou Ave., 102 50 ATHENS PROCUREMENT SECTION tel.: 2103202153, fax: ……………. 0100003 HEAD OFFICE HUMAN ATHENS CENTRE 21, El. Venizelou Ave., 102 50 ATHENS RESOURCES SECTION tel.: 2103202090, fax: 2103203961 0100004 HEAD OFFICE DOCUMENT ATHENS CENTRE 21, El. Venizelou Ave., 102 50 ATHENS MANAGEMENT TELEGRAMS AND TEST KEYS SECTION tel.: 2103202198, fax: …… 0100005 HEAD OFFICE PAYROLL ATHENS CENTRE 21, El. Venizelou Ave., 102 50 ATHENS MANAGEMENT SECTION tel.: 2103202128, fax: 2103236930 0100007 HEAD OFFICE SECURITY ATHENS CENTRE 21, El. Venizelou Ave., 102 50 ATHENS SECTION tel.: 2103202101, fax: 210 3202928 0100008 HEAD OFFICE SYSTEMIC CREDIT ATHENS CENTRE 3, Amerikis, 102 50 ATHENS INSTITUTIONS SUPERVISION SECTION A tel.: 2103205193, fax: …… 0100009 HEAD OFFICE BOOK ENTRY ATHENS CENTRE 21, El. Venizelou Ave., 102 50 ATHENS SECURITIES MANAGEMENT SECTION tel.: 2103202627, fax: 2103235747 0100010 HEAD OFFICE ARCHIVES ATHENS CENTRE 21, El. Venizelou Ave., 102 50 ATHENS SECTION tel.: 2103202206, fax: 2103203950 0100012 HEAD OFFICE RESERVES ATHENS CENTRE 21, El. Venizelou Ave., 102 50 ATHENS MANAGEMENT BACK UP SECTION tel.: 2103203766, fax: 2103220140 0100013 HEAD OFFICE FOREIGN ATHENS CENTRE 21, El. Venizelou Ave., 102 50 ATHENS EXCHANGE TRANSACTIONS SECTION tel.: 2103202895, fax: 2103236746 0100014 HEAD OFFICE SYSTEMIC CREDIT ATHENS CENTRE 3, Amerikis, 102 50 ATHENS INSTITUTIONS SUPERVISION SECTION B tel.: 2103205041, fax: …… 0100015 HEAD OFFICE PAYMENT ATHENS CENTRE 3, Amerikis, 102 50 ATHENS SYSTEMS OVERSIGHT SECTION tel.: 2103205073, fax: …… 0100016 HEAD OFFICE ESCB PROJECTS CHALANDRI 341, Mesogeion Ave., 152 31 CHALANDRI AUDIT SECTION tel.: 2106799748, fax: 2106799713 0100017 HEAD OFFICE DOCUMENTARY ATHENS CENTRE 21, El. -

Hazardous Waste Removal Locations for Eastern/Southern Europe and Israel Contract
Defense Logistics Agency Disposition Services Attachment 3 – Hazardous Waste Removal Locations For Eastern/Southern Europe and Israel Contract 1. Bulgaria 1.1. Novo Selo Training Area (NSTA) Kotel, Mokren Main Road I-7, Bulgaria 8994, Building 7005, 7006, 7016 2. Greece 1.1. Souda Bay NSA (Crete) Naval Support Activity Naval Base Souda Bay Supply Dept Bldg 6, Mouzouras Road, Chania, Crete 73100, Greece 3. Georgia Republic 1.1. Senaki Air Base Samegrelo and Zemo Svaneti Province, Senaki Airfield (Must contact Generator for address/coordinates) 4. Hungary 1.1. Papa Air Base Pápa, Vaszari út, Hanger-1, 8500 Hungary, Warehouse-1 1.2. Szolnok Air Base Szolnok, Kilián György utca 1, 5008 Hungary, Warehouse-2 1.3. Taborfalva Training Area Taborfalva Military Training Area, 2381, Taborfalva, Hungary 1.4. Várpalota-Veszprém Central Training Area (CTA) Hajmasker Rd, Maintenance Bay-100, 8100, Varpalota, Hungary 5. Israel 1.1. Site-1 (Must contact DLA COR, Generator or email USARMY Wiesbaden USAREUR Mailbox Environmental [email protected] 1.2. Site-2 (Must contact DLA COR, Generator or USARMY Wiesbaden USAREUR Mailbox Environmental [email protected] 6. Kosovo 1.1 Camp Bondsteel, Building-2033, APO AE 09340/UNIT, Camp Bondsteel 40148, 7. Poland 1.1. Bemowo Piskie Training Area (BPTA) Wojciecha Ketrznskiego, 12-230, Bemowo Piskie, Poland, White Maintenance Tent 1.2. Boleslawiec Aleja Wojska Polskiego 54, 59-700 Boleslawiec, Poland 1.3. Camp Karliki Camp Karliki road, Vehicle Maintenance bay 2, 68-100 Zagan, Poland 1.4. Drawsko Pomorskie - Konotop Barracks (DPTA) Nowy Konotop, 78-500 Drawsko Pomorskie, Poland, Hazardous Waste Storage Area Building-1000 1.5. -
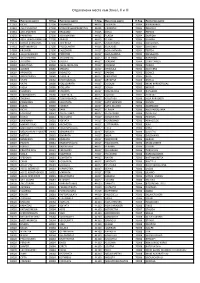
Print Sheet Remote1
Отдалечени места към Зона I, II и III П.Код Населено място П.Код Населено място П.Код Населено място П.Код Населено място 11361 KICELI 27100 KERAMIDIA 44015 LAGKADA 70004 KSEROKABOS 12351 AGIA VARVARA 27100 MONI FRAGKOPIDIMATOS 44015 LIKORRAXI 70004 PERVOLA 13561 AGII ANARGIRI 27100 TRAGANO 44015 OKSIA 70004 PEFKOS 13672 PARNITHA 27200 AGIA MARINA 44015 PLAGIA 70004 SKAFIDIA 13679 AGIA TRIADA PARNITHAS 27200 ANALICI 44015 PLIKATI 70004 STAFRIA 13679 KSENIA PARNITHAS 27200 ASTEREIKA 44015 PIRSOGIANNI 70004 SIKOLOGOS 14451 METAMORFOSI 27200 PALEOLANTHI 44015 XIONADES 70004 SINDONIA 14568 KRIONERI 27200 PALEOXORI 44017 AGIA VARVARA 70004 TERTSA 15342 AGIA PARASKEFI 27200 PERISTERI 44017 AGIA MARINA 70004 FAFLAGKOS 18010 AGIA MARINA 27300 AGIA MAFRA 44017 VEDERIKOS 70004 XONDROS 18010 AGKISTRI 27300 KALIVIA 44017 VERENIKI 70004 CARI FORADA 18010 EGINITISSA 28080 AGIOS NIKOLAOS 44017 VROSINA 70005 AVDOU 18010 ALONES 28080 GRIZATA 44017 VRISOULA 70005 ANO KERA 18010 APONISOS 28080 DIGALETO 44017 GARDIKI 70005 GONIES 18010 APOSPORIDES 28080 ZERVATA 44017 GKRIBOVO 70005 KERA 18010 VATHI 28080 KARAVOMILOS 44017 GRANITSA 70005 KRASIO 18010 VATHI 28080 KOULOURATA 44017 DIXOUNI 70005 MONI KARDIOTISSAS 18010 VIGLA 28080 POULATA 44017 DOVLA 70005 MOXOS 18010 VLAXIDES 28080 STAVERIS 44017 DOMOLESSA 70005 POTAMIES 18010 GIANNAKIDES 28080 TZANEKATA 44017 ZALOGO 70005 SFENDILI 18010 THERMA 28080 TSAKARISIANOS 44017 KALLITHEA 70006 AGIA PARASKEFI 18010 KANAKIDES 28080 XALIOTATA 44017 KATO VERENIKI 70006 AGNOS 18010 KLIMA 28080 XARAKTI 44017 KATO ZALOGO -

List of Bank Branches by Branch Name 31/12/2014
LIST OF BANK BRANCHES BY BRANCH NAME 31/12/2014 PIRAEUS BANK S.A. BRANCH NAMEHEBIC AREA ADDRESS TELEPHONE NUMBER / FAX 1866 SQUARE, CHANIA0172758 CHANIA 70, Chatzimichali Giannari str., 731 35 CHANIA tel.: 2821029100, fax: 2821050710 25TH AVGOUSTOU, HERAKLIO0172751 HERAKLEIO 39, 25th Avgoustou str., 712 02 HERAKLEIO tel.: 2810247610, fax: 2810228324 3RD SEPTEMVRIOU, OMONIA 0172039 ATHENS 28, 3rd Septemvriou & Kapodistriou str., 104 32 ATHENS BRANCH tel.: 2105205100, fax: 2105235138 62 MARTIRON HERAKLIOU0172761 HERAKLEIO 97, 62 Martiron Ave. & 1, Dionysiou Fragiadaki str., 713 04 HERAKLEIO tel.: 2810264330, fax: 2810255997 ABELONAS0171393 AMPELONAS 1, Goltsiou & 5, Thermopylon str., 404 00 AMPELONAS tel.: 2492306000, fax: 2492031402 ACHARNON0172058 ATHENS 421, Acharnon str., 111 43 ATHENS tel.: 2102588070, fax: 2102520490 ACHARNON STR.0171592 ATHENS 205, Acharnon & Niovis str.s., 104 46 ATHENS tel.: 2166008310, fax: 2168003817 AEGINA BRANCH0172124 AEGINA Dimokratias Coastal Ave. & Leonardou Lada str., 180 10 AEGINA tel.: 2297029890, fax: 2297028882 AG. ACHILLIOU0171563 LARISSA 146, El. Venizelou str., 412 22 LARISSA tel.: 2416000160, fax: 2416007111 AG. ANTONIOU, PERISTERI0172048 PERISTERI 33, Ethnarchou Makariou str., 121 31 PERISTERI tel.: 2105783020, fax: 2105773224 AG. DIMITRIOS0171555 AGIOS DIMITRIOS 266, Vouliagmenis Ave., 173 43 AGIOS DIMITRIOS tel.: 2160005020, fax: 2109765714 AG. DIMITRIOU0172085 AGIOS DIMITRIOS 173, Ag. Dimitriou Ave., 173 43 AGIOS DIMITRIOS tel.: 2109758170, fax: 2109764036 AG. DIMITRIOU, THESSALONIKI0172203 THESSALONIKI 77, Ag. Dimitriou str., 546 33 THESSALONIKI tel.: 2310254860, fax: 2310223843 AG. IOANNI RENTI SQ. BRANCH0172136 AG. I. RENTIS 4, Konstantinou Palaiologou str., Ag. I. Renti sq., 182 33 AG. I. RENTIS tel.: 2104838120, fax: 2104832389 AG. IOANNOU, AG. PARASKEVI0172010 AGIA PARASKEVI 40, Ag. Ioannou str., 153 42 AGIA PARASKEVI tel.: 2106085800, fax: 2106011128 AG. -

Guidance Note CVED for US Military Basis.Pdf
Guidance note for the fulfilment of CVED-Ps for the non-confirming consignments destined to US military basis located in Italy, Germany, Greece and Spain Based on agreement with US Public Health Command for Europe region, the US military basis located in Italy, Germany, Greece and Spain will start to use TRACES for the notification of the arrival of the goods in their basis to the entry EU border inspection post as required for the consignments in transit. To allow access for US Military basis for the CVEDS accompanying the consignments in TRACES to sign the CVED Control part, the following rules need to be respected. 1. In Box 8 “Delivery address” the delivery address in US military basis should be in accordance with the entries listed in attached Annex under the column “Delivery Name & Address”. 2. In box 18 “For transit to 3rd Country” the exit BIP should be entry listed in attached Annex under the column “US Base/exit BIP ID”. Example: The consignment delivery address is in Ramstein Flugplatz as Central meat Processing Plant (CMPP), Geb. 1200. This delivery address is indicated in Box 8, and according to the correspondence table, in box 18. The exit BIP should be selected as US-DE-Ramstein-1, and the destination third country as United States: Annex Correspondence of US military base delivery addresses with the exit EU border inspection posts selection in TRACES US Base/exit BIP TRACES unit ID code Delivery Name & Address Approval number US-DE-Ramstein-1 DEUS101 Central Meat Processing Plant (CMPP), US-DE-Ramstein-1 Geb. -

Tower-Mansions of Crete. a Multidisciplinary Approach to Learn Built Heritage
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by PORTO Publications Open Repository TOrino Politecnico di Torino Porto Institutional Repository [Proceeding] Tower-mansions of Crete. A multidisciplinary approach to learn built heritage Original Citation: Maglio Emma (2016). Tower-mansions of Crete. A multidisciplinary approach to learn built heritage. In: FORTMED 2016. pp. 487-494 Availability: This version is available at : http://porto.polito.it/2675116/ since: June 2017 Publisher: DidaPress Terms of use: This article is made available under terms and conditions applicable to Open Access Policy Article ("Public - All rights reserved") , as described at http://porto.polito.it/terms_and_conditions. html Porto, the institutional repository of the Politecnico di Torino, is provided by the University Library and the IT-Services. The aim is to enable open access to all the world. Please share with us how this access benefits you. Your story matters. (Article begins on next page) Defensive Architecture of the MeDiterrAneAn 3 Xv to Xviii centuries Giorgio verDiAni (ed.) PROCEEDINGS of the International Conference on Modern Age Fortifications of the Mediterranean Coast FORTMED 2016 DEFENSIVE ARCHITECTURE OF THE MEDITERRANEAN XV TO XVIII CENTURIES Vol. III Editor Giorgio Verdiani Università degli Studi di Firenze Dipartimento di Architettura PUBLISHED BY DIDAPRESS FORTMED 2016, FIRENZE Atti del Congresso / Conference Proceedings / Colección Congresos UNIFI Tutti i contenuti della presente pubblicazione sono stati soggetti a revisione da parte del Comitato Scientifico di FORTMED 2016, secondo il processo della “peer review”. All the contents of this book has been reviewed by the FORTMED 2016 Scientific Committee according to the “peer review” process. -

List of Bank Branches by Area 31/03/2021
LIST OF BANK BRANCHES BY AREA 31/03/2021 PIRAEUS BANK S.A. AREA / LOCATIONHEBIC BRANCH NAME ADDRESS TELEPHONE NUMBER / FAX NUMBER AEGINA / Aegina 0172124 AEGINA BRANCH Dimokratias Coastal Ave. & Leonardou Lada str., 180 10 AEGINA tel.: 2297029890, fax: 2297028882 AGIA / Agia 0171249 AGIAS 33, 25th Martiou str., 400 03 AGIA tel.: 2494024530, fax: 2494023988 AGIA MARINA / Agia Marina, Platanias 0171635 AG. MARINA CHANIA BRANCH 9th klm, Chania-Kissamos National Rd., 730 14 AGIA MARINA Chania tel.: 2821036650, fax: 2821068079 AGIA PARASKEVI / Ag. Paraskevi 0171561 AG. PARASKEVI 507, Mesogion Ave., 153 43 AGIA PARASKEVI tel.: 2168008300, fax: 2106006426 AGIA PARASKEVI / Agia Paraskevi Square 0172009 AG. PARASKEVIS 406, Mesogion Ave. & 2 Iroon Politexniou, 153 42 AGIA PARASKEVI tel.: 2106085820, fax: 2106009558 AGIA PARASKEVI / Agiou Ioannou 0172010 AG. IOANNOU, AG. PARASKEVI 40, Ag. Ioannou str., 153 42 AGIA PARASKEVI tel.: 2106085800, fax: 2106011128 AGIOI ANARGYROI / Agioi Anargyroi 0172143 AGIOI ANARGYRI BRANCH 31, Agion Anargyron & Kefallinias str., 135 61 AGIOI ANARGYROI tel.: 2102694060, fax: 2102696096 AGIOS DIMITRIOS / Ag. Dimitrios 0171555 AG. DIMITRIOS 266, Vouliagmenis Ave., 173 43 AGIOS DIMITRIOS tel.: 2160005020, fax: 2109765714 AGIOS DIMITRIOS / Agios Dimitrios 0171038 AGIOS DIMITRIOS 1-3, Theodorou Kolokotroni str., 173 43 AGIOS DIMITRIOS tel.: 2164002150, fax: 2167007112 AGIOS DIMITRIOS / Agios Dimitrios 0172193 L. VOULIAGMENIS 380 380, Vouliagmenis Ave. & 2, Spirou Miliou & Dodekanisou str., 173 42 BRANCH AGIOS DIMITRIOS tel.: 2109903000, fax: 2109902075, 2109902745 AGIOS KIRYKOS / Agios Kirykos 0171150 AGIOS KIRIKOS IKARIAS Agios Kirykos, 833 00 AGIOS KIRYKOS tel.: 2275025040, fax: 2275023193 AGIOS NIKOLAOS / Agios Nikolaos 0172756 AG. NIKOLAOU, CRETE 18, Mich. Sfakianaki str., 721 00 AGIOS NIKOLAOS tel.: 2841086000, fax: 2841025609 AGIOS STEFANOS / Agios Stefanos 0172083 AG. -

Greek Commercial Banks by Total Assets 31.12.2015
LIST OF BANK BRANCHES BY AREA 30/06/2021 PIRAEUS BANK S.A. AREA / LOCATIONHEBIC BRANCH NAME ADDRESS TELEPHONE NUMBER / FAX NUMBER AEGINA / Aegina 0172124 AEGINA BRANCH Dimokratias Coastal Ave. & Leonardou Lada str., 180 10 AEGINA tel.: 2297029890, fax: 2297028882 AGIA / Agia 0171249 AGIAS 33, 25th Martiou str., 400 03 AGIA tel.: 2494024530, fax: 2494023988 AGIA MARINA / Agia Marina, Platanias 0171635 AG. MARINA CHANIA BRANCH 9th klm, Chania-Kissamos National Rd., 730 14 AGIA MARINA Chania tel.: 2821036650, fax: 2821068079 AGIA PARASKEVI / Ag. Paraskevi 0171561 AG. PARASKEVI 507, Mesogion Ave., 153 43 AGIA PARASKEVI tel.: 2168008300, fax: 2106006426 AGIA PARASKEVI / Agia Paraskevi Square0172009 AG. PARASKEVIS 406, Mesogion Ave. & 2 Iroon Politexniou, 153 42 AGIA PARASKEVI tel.: 2106085820, fax: 2106009558 AGIA PARASKEVI / Agiou Ioannou 0172010 AG. IOANNOU, AG. PARASKEVI40, Ag. Ioannou str., 153 42 AGIA PARASKEVI tel.: 2106085800, fax: 2106011128 AGIOI ANARGYROI / Agioi Anargyroi 0172143 AGIOI ANARGYRI BRANCH 31, Agion Anargyron & Kefallinias str., 135 61 AGIOI ANARGYROI tel.: 2102694060, fax: 2102696096 AGIOS DIMITRIOS / Ag. Dimitrios 0171555 AG. DIMITRIOS 266, Vouliagmenis Ave., 173 43 AGIOS DIMITRIOS tel.: 2160005020, fax: 2109765714 AGIOS DIMITRIOS / Agios Dimitrios 0171038 AGIOS DIMITRIOS 1-3, Theodorou Kolokotroni str., 173 43 AGIOS DIMITRIOS tel.: 2164002150, fax: 2167007112 AGIOS DIMITRIOS / Agios Dimitrios 0172193 L. VOULIAGMENIS 380 380, Vouliagmenis Ave. & 2, Spirou Miliou & Dodekanisou str., 173 42 BRANCH AGIOS DIMITRIOS tel.: 2109903000, fax: 2109902075, 2109902745 AGIOS KIRYKOS / Agios Kirykos 0171150 AGIOS KIRIKOS IKARIAS Agios Kirykos, 833 00 AGIOS KIRYKOS tel.: 2275025040, fax: 2275023193 AGIOS NIKOLAOS / Agios Nikolaos 0172756 AG. NIKOLAOU, CRETE 18, Mich. Sfakianaki str., 721 00 AGIOS NIKOLAOS tel.: 2841086000, fax: 2841025609 AGIOS STEFANOS / Agios Stefanos 0172083 AG. -

CRETE BIRD REPORT 1998 Compiled by Denis Townsend Edited By: Stephanie Coghlan
CRETE BIRD REPORT 1998 compiled by Denis Townsend edited by: Stephanie Coghlan. CRETE BIRD REPORT 1998 Introduction to Crete Bird Report 1998 Thank you to all those who have spared time to send in reports of their sightings. It is all helping to give us a better picture of the birds on the island of Crete. Denis Townsend has worked hard collating all the records you have sent in. Many thanks again to him for all his hard work without which this report would not appear. Please help him by submitting any further records in the order used in this report. Any records of weather and other reports of mammals, butterflies and moths and amphibians and reptiles recorded, preferably with sites are also very useful. Weather: The winter weather was mild until early May when there was a period of cool wet weather but then the summer settled down to the usual hot dry season. The Autumn and Winter did not have any exceptional weather which affected bird numbers. Names and spelling: Many more place names are appearing in the text and we have refer you to the Nelles Verlag 1: 200,000 which hopefully agree with the spelling we have used. Illustrations: Many thanks to Ken Baldridge for his illustrations which we have used in this report. The Bird List follows the order in “The Birds of Greece” Handrinos/Akriotis.1997. R – resident, PM - passage migrant. AV- accidental vagrant, V- vagrant , SV-summer visitor, WV- winter visitor, STATUS: rare, scarce, common, locally common, introduced. LITTLE GREBE Tachybaptus ruficollis R PM Winter -Spring : max 35 Bramiana reservoir 1.1. -

Bips Guidance Transit Tranship
EUROPEAN COMMISSION DIRECTORATE-GENERAL FOR HEALTH AND FOOD SAFETY Food chain: stakeholder and international relations Multilateral International relations SANCO/10844/2011 REVISION 4, 01.12.2016 EN WORKING DOCUMENT GENERAL GUIDANCE FOR CONSIGNMENTS OF LIVE ANIMALS AND ANIMAL PRODUCTS FROM THIRD COUNTRIES IN TRANSIT OR TRANSHIPMENT This document has been established for information purposes only. It has not been adopted or in any way approved by the European Commission. The European Commission does not guarantee the accuracy of the information provided, nor does it accept responsibility for any use made thereof. Users should therefore take all necessary precautions before using this information, which they use entirely at their own risk. PURPOSE OF THIS DOCUMENT This document is mainly directed at competent authorities and more specifically at border inspection posts, and aims to give guidance on the implementation of the requirements governing the import and transit control system for live animals and animal products originating from third countries. NOTE This document is an evolving document and may be updated as necessary to take account of experiences and information from competent authorities and private operators in third countries, from importers and from the Commission's inspection service, the Food and Veterinary Office (FVO). i ABBREVIATIONS AND DEFINITIONS USED IN THE GUIDANCE DOCUMENT BIP Border Inspection Post as defined in Council Directives 91/496/EEC and 97/78/EC CVED Common Veterinary Entry Document for animal products