Jennifer Galloway's 2010 Paper
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

British Columbia Regional Guide Cat
National Marine Weather Guide British Columbia Regional Guide Cat. No. En56-240/3-2015E-PDF 978-1-100-25953-6 Terms of Usage Information contained in this publication or product may be reproduced, in part or in whole, and by any means, for personal or public non-commercial purposes, without charge or further permission, unless otherwise specified. You are asked to: • Exercise due diligence in ensuring the accuracy of the materials reproduced; • Indicate both the complete title of the materials reproduced, as well as the author organization; and • Indicate that the reproduction is a copy of an official work that is published by the Government of Canada and that the reproduction has not been produced in affiliation with or with the endorsement of the Government of Canada. Commercial reproduction and distribution is prohibited except with written permission from the author. For more information, please contact Environment Canada’s Inquiry Centre at 1-800-668-6767 (in Canada only) or 819-997-2800 or email to [email protected]. Disclaimer: Her Majesty is not responsible for the accuracy or completeness of the information contained in the reproduced material. Her Majesty shall at all times be indemnified and held harmless against any and all claims whatsoever arising out of negligence or other fault in the use of the information contained in this publication or product. Photo credits Cover Left: Chris Gibbons Cover Center: Chris Gibbons Cover Right: Ed Goski Page I: Ed Goski Page II: top left - Chris Gibbons, top right - Matt MacDonald, bottom - André Besson Page VI: Chris Gibbons Page 1: Chris Gibbons Page 5: Lisa West Page 8: Matt MacDonald Page 13: André Besson Page 15: Chris Gibbons Page 42: Lisa West Page 49: Chris Gibbons Page 119: Lisa West Page 138: Matt MacDonald Page 142: Matt MacDonald Acknowledgments Without the works of Owen Lange, this chapter would not have been possible. -

3LMANUSCRIPT REPORT SERIES No. 36
DFO - L bra y MPOBibio heque II 1 111111 11 11 11 V I 1 120235441 3LMANUSCRIPT REPORT SERIES No. 36 Some If:eat/viz& 3,5,unamia, Olt the Yacific ettadt of South and ✓ cuith anwitica, T. S. Murty, S. 0. Wigen and R. Chawla Marine Sciences Directorate 975 Department of the Environment, Ottawa Marine Sciences Directorate Manuscript. Report Series No. 36 SOME FEATURES OF TSUNAMIS ON THE PACIFIC COAST OF SOUTH AND NORTH AM ERICA . 5 . Molly S . O. Wigen and R. Chawla 1975 Published by Publie par Environment Environnement Canada Canada I' Fisheries and Service des !Aches Marine Service et des sciences de la mer Office of the Editor Bureau du fiedacteur 116 Lisgar, Ottawa K1 A Of13 1 Preface This paper is to be published in Spanish in the Proceedings of the Tsunami Committee XVII Meeting, Lima, Peru 20-31 Aug. 1973, under the International Association of Seismology and Physics of the Earth Interior. 2 Table of Contents Page Abstract - Resume 5 1. Introduction 7 2. Resonance characteristics of sonic inlets on the Pacific Coast of Soulh and North America 13 3. Secondary undulations 25 4. Tsunami forerunner 33 5. Initial withdrawal of water 33 6. Conclusions 35 7. References 37 3 4 i Abstract In order to investigate the response of inlets to tsunamis, the resonance characteristics of some inlets on the coast of Chile have been deduced through simple analytical considerations. A comparison is made with the inlets of southeast Alaska, the mainland coast of British Columbia and Vancouver Island. It is shown that the general level of intensif yy of secondary undulations is highest for Vancouver Island inlets, and least for those of Chile and Alaska. -

Northisle Copper and Gold Inc
Ministry of Energy, Mines & Petroleum Resources Mining & Minerals Division Assessment Report BC Geological Survey Title Page and Summary TYPE OF REPORT [type of survey(s)]: Archaeological Impact Assessment TOTAL COST: $14,254 AUTHOR(S): Morgan Bartlett, Robbin Chatan, Konstantin Lesnikov SIGNATURE(S): NOTICE OF WORK PERMIT NUMBER(S)/DATE(S): MX-8-271 / December 22, 2011 YEAR OF WORK: 2012 STATEMENT OF WORK - CASH PAYMENTS EVENT NUMBER(S)/DATE(S): 5421553 PROPERTY NAME: Island Copper East Block CLAIM NAME(S) (on which the work was done): Mo 4, Mo 5 COMMODITIES SOUGHT: N/A MINERAL INVENTORY MINFILE NUMBER(S), IF KNOWN: 092L 273 MINING DIVISION: Nanaimo NTS/BCGS: 092L/11 / 92L.054 o '" o '" LATITUDE: -127 22 21 LONGITUDE: 50 35 18 (at centre of work) OWNER(S): 1) NorthIsle Copper and Gold Inc. 2) MAILING ADDRESS: #2050-1111 West Georgia Street Vancouver BC V6E 4M3 OPERATOR(S) [who paid for the work]: 1) 2) MAILING ADDRESS: PROPERTY GEOLOGY KEYWORDS (lithology, age, stratigraphy, structure, alteration, mineralization, size and attitude): Bonanza Volcanics, Quatsino Formation, Karmutsen Formation, Island Plutonic Suite, porphyritic granitoid rocks, granodiorite, andesite, basalt, limestone, Upper Triassic, middle Jurassic, Island Copper Mine REFERENCES TO PREVIOUS ASSESSMENT WORK AND ASSESSMENT REPORT NUMBERS: AR 2659, 1681, 14393, 15884, 15707, 11460, 15024, 15077, 15367, 16510, 15024, 15077, 15367, 16510, 17368, 32722 Next Page TYPE OF WORK IN EXTENT OF WORK ON WHICH CLAIMS PROJECT COSTS THIS REPORT (IN METRIC UNITS) APPORTIONED (incl. support) -
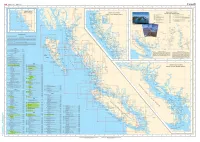
CHS Index Chart
Fisheries and Oceans Pi'lches et Oceans Canada Canada Canada ••• 137 ° 136 " 134" 133 ° 131° 129 ° ,.. 126 ° 125 " 124 " 123° 122° 119° 124 ° 118 ° GENERAL CHARTS CARTES GENERALES SMALL-CRAFT CHARTS REV ILLAG IGEDO LARGER SCALE CHARTS ISLAND CARTES POUR EMBARCATIONS CARTES A PLUS GRANDE ECHELLE 3050 Kootenay Lake and Rovet 75 000 3311 Sunshtno Coast- Vancouver Ha rbour lo/A 3052 Okanagan Lako so 000 Desolatoon Sound 40 000 3053 Shuswap Lake so 000 331:2 JerviS Intel ond/et Do•o latoon Sound 0 305S Waneta to /~ Hugh Keen leyside Dam 20 000 Vo,ous Scolo•JEche tle• vo"h• > z 3056 Hugh Koon loySido Dam to/A Burlon 40 000 3313 Gull Islands and Ad jacent Watotways/el les Vo1es Navigables Ad1acentes ~ ' 3057 Button IO/~ 1\rrowhood 40 000 Variou• Scale•/Echel le• vo"~"' 3058 Arrowhead lo/6 Rovo lotoko Go «m• 20 000 3488 Fro5er River/F I&uve Fraser, Cre•<ent l5land 3061 " "'"'on Lake and/ol Hamson R1ver '""'' to/~ Hon loon Mills 20 000 Harrison Lake 40 000 3469 Fraser Rovor/Fiouve Fr8set, Pattullo B"dgo Harrison R1ver 30 000 to/a Crescent Is land 20 000 Pitt River and/ot Poll l ak e 25 000 Stuaot L a~e (Not•howniP••rnd•qu o!) 50 000 54 " ~f--- ' ~ 0 "" I < ''"0' 't)Go iUn "?1- Cocoov• 3053 Foo ..o<o o l 0 "' GJ ,. Shu wap .•. Lake CANAOA !'; "'""""'' •·o~ d 130" 125° 120 " •5"omouo .,cocho Cceo> ,... ,. ' GJ ... ' <om l oops ~ DIXON E'N TRANCE' LEGENO/LEGENDE • •• • Scales smaller ttlan 1"40 000 Ectlellas plus petites qua 1:40 000 '' GJ Scales 1:40 000 and larger Ectlelles 1: 40 000 et plus grande& CHART SCALE Chart soale os the rat10 of one umt of d1stance 011 the cha" to the actual d.stance on the Earth's surface expressed on tho same unots. -

Climate Change and Decadal to Centennial-Scale Periodicities Recorded in a Late Holocene NE Pacific Marine Record: Examining the Role of Solar Forcing
Palaeogeography, Palaeoclimatology, Palaeoecology 386 (2013) 669–689 Contents lists available at SciVerse ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Climate change and decadal to centennial-scale periodicities recorded in a late Holocene NE Pacific marine record: Examining the role of solar forcing J.M. Galloway a,⁎, A. Wigston a, R.T. Patterson a, G.T. Swindles b, E. Reinhardt c, H.M. Roe d a Department of Earth Sciences and Ottawa Geoscience Centre, Carleton University, 1125 Colonel By Drive, Ottawa, Ontario K1S 5B6, Canada b School of Geography, University of Leeds, Leeds LS2 9JT, United Kingdom c School of Geography and Earth Sciences, McMaster University, Hamilton, ON L8S 4K1, Canada d School of Geography, Archaeology and Palaeoecology, Queen's University Belfast, Belfast BT7 1NN, United Kingdom article info abstract Article history: We present a decadal-scale late Holocene climate record based on diatoms, biogenic silica, and grain size Received 29 November 2012 from a 12-m sediment core (VEC02A04) obtained from Frederick Sound in the Seymour-Belize Inlet Complex Received in revised form 21 June 2013 of British Columbia, Canada. Sediments are characterized by graded, massive, and laminated intervals. Accepted 22 June 2013 Laminated intervals are most common between c. 2948–2708 cal. yr BP and c. 1992–1727 cal. yr BP. Available online 4 July 2013 Increased preservation of laminated sediments and diatom assemblage changes at this time suggest that cli- mate became moderately drier and cooler relative to the preceding and succeeding intervals. Spectral and Keywords: fi Late Holocene wavelet analyses are used to test for statistically signi cant periodicities in time series of proxies of primary Climate change production (total diatom abundance, biogenic silica) and hydrology (grain size) preserved in the Frederick Diatoms Sound record. -

REC-EWED by APR 1 1,2009 J
PROSPECTING ASSESSMENT REPORT ON THE SEYMOUR INLET PROJECT (WIGWAM CLAIMS, Tenure # 572620 & 572621) (Pt/Os/Rh/lr ALASKAN TYPE INTRUSION) Seymour Inlet, Vancouver Mining Division NTS 92 M/2 (92M.017) Latitude 51°08'19", Longitude: 126°43'48" REC-EWED By APR 1 1,2009 J. T. ~Ihearer, M.Sc., P.Geo. Gold Commissioner's MfM5' - 2330 Tyner Street VANCOUVER, B.C. Port Coquitlam, BC !fJ r V3C 221 ;;e; ~~ • Phone:604-970-6402 ~ 1:1 I, ,j E-mail: [email protected] ~ [,! ,1 ~ri' " ("-I For § [~, l;j /2,. , i &-1' ...; ~,. "< ", Homegold Resources ltd /O-'"iI! c.":'/ v c' Unit 5 - 2330 Tyner Street t: £; ~ c«( Port Coquitlam, BC v c .. ~';.".;;- "'" V3C 221 ~J September 15, 2008 Fieldwork completed between August 1, 2008 and August 30, 2008 r- • TABLE of CONTENTS PAGE INTRODUCTION 1 PROPERTy 2 LOCATION, ACCESS and TOPOGRAPHY : 3 HISTORY and DEVELOPMENT 4 REGIONAL GEOLOGY 5 LOCAL GEOLOGy 6 ROCK DESCRIPTIONS 7 2008 PROSPECTING and GEOLOGY 9 CONCLUSIONS and RECOMMENDATIONS 10 ESTIMATE of COSTS for FUTURE WORK 11 STATEMENT of QUALIFICATIONS 12 REFERENCES 13 APPENDICES Appendix I Statement of Costs 14 Appendix II List of Specimens 15 Appendix III Titaniferous Zoned Pt-OZ-Rh/lr Vadadium Deposit Model And Magmatic Oxide Deposit Model 17 Prospecting Report on the Seymour Inlet Project (Wigwam Claims) September 15, 2008 Page ii • LIST of ILLUSTRATIONS and TABLES Following Page FIGURE 1 Property Location, 1":190km ii FIGURE 2 Detail Property Location 1 FIGURE 3 Claim Location Map, 1:51,000 2 FIGURE 4 Regional Geology Map, 1:147,869 4 FIGURE 5 Local Geology, 1:50,000 5 FIGURE 6 Local Prospecting Map, 1:10,000, Rainbow Creek Area 8 FIGURE 7 Local Prospecting Map, 1:10,000, Haig Bay Area 9 TABLES Page TABLE I List of Claims ! •••••••••••• 2 Prospecting Report on the Seymour Inlet Project (Wigwam Claims) September 15, 2008 Page iii ----------------~-------- pW INTRODUCTION The Wigwam Project is located near the head of Seymour Inlet, BC, a narrow coastal mainland fjord 85km east of the north tip of Vancouver Island in the Vancouver Mining Division. -

Postglacial Vegetation and Climate Dynamics in the Seymour-Belize Inlet Complex, Central Coastal British Columbia, Canada: Palynological Evidence from Tiny Lake
Postglacial vegetation and climate dynamics in the Seymour-Belize Inlet Complex, central coastal British Columbia, Canada: palynological evidence from Tiny Lake. Galloway, J. M., Doherty, C. T., Patterson, R. T., & Roe, H. (2009). Postglacial vegetation and climate dynamics in the Seymour-Belize Inlet Complex, central coastal British Columbia, Canada: palynological evidence from Tiny Lake. Journal of Quaternary Science, 24(4), 322-335. https://doi.org/10.1002/jqs.1232 Published in: Journal of Quaternary Science Document Version: Early version, also known as pre-print Queen's University Belfast - Research Portal: Link to publication record in Queen's University Belfast Research Portal General rights Copyright for the publications made accessible via the Queen's University Belfast Research Portal is retained by the author(s) and / or other copyright owners and it is a condition of accessing these publications that users recognise and abide by the legal requirements associated with these rights. Take down policy The Research Portal is Queen's institutional repository that provides access to Queen's research output. Every effort has been made to ensure that content in the Research Portal does not infringe any person's rights, or applicable UK laws. If you discover content in the Research Portal that you believe breaches copyright or violates any law, please contact [email protected]. Download date:23. Sep. 2021 JOURNAL OF QUATERNARY SCIENCE (2009) 24(4) 322–335 Copyright ß 2008 John Wiley & Sons, Ltd. Published online 23 October 2008 in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/jqs.1232 Postglacial vegetation and climate dynamics in the Seymour-Belize Inlet Complex, central coastal British Columbia, Canada: palynological evidence from Tiny Lake JENNIFER M. -

Scale Site SS Region SS District Site Name SS Location Phone
Scale SS SS Site Region District Site Name SS Location Phone 001 RCB DQU MISC SITES SIFR 01B RWC DQC ABFAM TEMP SITE SAME AS 1BB 2505574201 1001 ROM DPG BKB CEDAR Road past 4G3 on the old Lamming Ce 2505690096 1002 ROM DPG JOHN DUNCAN RESIDENCE 7750 Lower Mud river Road. 1003 RWC DCR PROBYN LOG LTD. Located at WFP Menzies#1 Scale Site 1004 RWC DCR MATCHLEE LTD PARTNERSHIP Tsowwin River estuary Tahsis Inlet 2502872120 1005 RSK DND TOMPKINS POST AND RAIL Across the street from old corwood 1006 RWC DNI CANADIAN OVERSEAS FOG CREEK - North side of King Isla 6046820425 1007 RKB DSE DYNAMIC WOOD PRODUCTS 1839 Brilliant Road Castlegar BC 2503653669 1008 RWC DCR ROBERT (ANDY) ANDERSEN Mobile Scale Site for use in marine 1009 ROM DPG DUNKLEY- LEASE OF SITE 411 BEAR LAKE Winton Bear lake site- Current Leas 2509984421 101 RWC DNI WESTERN FOREST PRODUCTS INC. MAHATTA RIVER (Quatsino Sound) - Lo 2502863767 1010 RWC DCR WESTERN FOREST PRODUCTS INC. STAFFORD Stafford Lake , end of Loughborough 2502863767 1011 RWC DSI LADYSMITH WFP VIRTUAL WEIGH SCALE Latitude 48 59' 57.79"N 2507204200 1012 RWC DNI BELLA COOLA RESOURCE SOCIETY (Bella Coola Community Forest) VIRT 2509822515 1013 RWC DSI L AND Y CUTTING EDGE MILL The old Duncan Valley Timber site o 2507151678 1014 RWC DNI INTERNATIONAL FOREST PRODUCTS LTD Sandal Bay - Water Scale. 2 out of 2502861881 1015 RWC DCR BRUCE EDWARD REYNOLDS Mobile Scale Site for use in marine 1016 RWC DSI MUD BAY COASTLAND VIRTUAL W/S Ladysmith virtual site 2507541962 1017 RWC DSI MUD BAY COASTLAND VIRTUAL W/S Coastland Virtual Weigh Scale at Mu 2507541962 1018 RTO DOS NORTH ENDERBY TIMBER Malakwa Scales 2508389668 1019 RWC DSI HAULBACK MILLYARD GALIANO 200 Haulback Road, DL 14 Galiano Is 102 RWC DNI PORT MCNEILL PORT MCNEILL 2502863767 1020 RWC DSI KURUCZ ROVING Roving, Port Alberni area 1021 RWC DNI INTERNATIONAL FOREST PRODUCTS LTD-DEAN 1 Dean Channel Heli Water Scale. -

Late Pleistocene Palaeoenvironments, Archaeology, and Indicators of a Glacial Refugium on Northern Vancouver Island, Canada
Late Pleistocene palaeoenvironments, archaeology, and indicators of a glacial refugium on northern Vancouver Island, Canada by Christopher Franklin George Hebda B.A., University of Victoria, 2014 A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of MASTER OF ARTS in the Department of Anthropology © Christopher Franklin George Hebda, 2019 University of Victoria All rights reserved. This thesis may not be reproduced in whole or in part, by photocopy or other means, without the permission of the author. We acknowledge with respect the Lekwungen peoples on whose territory the university stands and the Songhees, Esquimalt and W̱ SÁNEĆ peoples whose historical relationships with the land continue to this day. ii Supervisory Committee Late Pleistocene palaeoenvironments, archaeology, and indicators of a glacial refugium on northern Vancouver Island, Canada by Christopher Franklin George Hebda B.A., University of Victoria, 2014 Supervisory Committee Dr. Quentin Mackie, Department of Anthropology Co-Supervisor Dr. Duncan McLaren, Department of Anthropology Co-Supervisor iii Abstract Recent research has revealed human settlement on the Pacific coast of Canada extending back nearly 14,000 years, but much of the late Pleistocene record is unknown due to shifting sea levels, poor understanding of Cordilleran ice extent, and limited research on the biota of the coast during this time. This study, undertaken in Quatsino First Nation and ‘Namgis First Nation territories as part of the Northern Vancouver Island Archaeology and Palaeoecology Project, employs modern multi-proxy analysis of lake sediment cores from two sites on northern Vancouver Island to reconstruct palaeoenvironments during and immediately following the Fraser Glaciation in coastal British Columbia. -

Inpfc Tag Recovery Data Format (All-Agency)
NPAFC Doc. 1026 Rev. Data Formats and Codes for the INPFC/NPAFC All-Agency High Seas Salmon and Steelhead Tag Recovery Computer Data File (1956-2005) by Katherine W. Myers and Robert V. Walker High Seas Salmon Research Program School of Aquatic and Fishery Sciences University of Washington Seattle, Washington, USA Submitted to the North Pacific Anadromous Fish Commission April 2007 THIS PAPER MAY BE CITED IN THE FOLLOWING MANNER: Myers, K. W., and R. V. Walker. 2007. Data formats and codes for the INPFC/NPAFC all- agency high seas salmon and steelhead tag recovery computer data file (1956-2005). NPAFC Doc. 1026. 26 pp. (Available at http://www.npafc.org). Data Formats and Codes for the INPFC/NPAFC All-Agency High Seas Salmon and Steelhead Tag Recovery Computer Data File (1956-2005) Katherine W. Myers and Robert V. Walker School of Aquatic and Fishery Sciences University of Washington Seattle, Washington, USA Abstract: This document lists the formats and codes used since 1971 for the all-agency INPFC- NPAFC high seas salmon and steelhead tag recovery computer data file (1956-2005). A brief overview and history of the computer data file is provided. Overview and Brief History of the High Seas Tag Recovery Data File This document lists the data formats and codes used since 1971 for the all-agency International North Pacific Fisheries Commission (INPFC) and North Pacific Anadromous Fish Commission (NPAFC) high seas salmon and steelhead high tag recovery computer data file (1956-2005; Tables 1- 3; Figs. 1-9). Harris (1989) briefly described the history of the development of the original INPFC all-agency high seas salmon and steelhead tag recovery computer data file, as follows: “Recoveries from high seas tagging experiments have been reported by the various INPFC national sections in the form of INPFC Documents. -

Oceanography of the British Columbia Coast
CANADIAN SPECIAL PUBLICATION OF FISHERIES AND AQUATIC SCIENCES 56 DFO - L bra y / MPO B bliothèque Oceanography RI II I 111 II I I II 12038889 of the British Columbia Coast Cover photograph West Coast Moresby Island by Dr. Pat McLaren, Pacific Geoscience Centre, Sidney, B.C. CANADIAN SPECIAL PUBLICATION OF FISHERIES AND AQUATIC SCIENCES 56 Oceanography of the British Columbia Coast RICHARD E. THOMSON Department of Fisheries and Oceans Ocean Physics Division Institute of Ocean Sciences Sidney, British Columbia DEPARTMENT OF FISHERIES AND OCEANS Ottawa 1981 ©Minister of Supply and Services Canada 1981 Available from authorized bookstore agents and other bookstores, or you may send your prepaid order to the Canadian Government Publishing Centre Supply and Service Canada, Hull, Que. K1A 0S9 Make cheques or money orders payable in Canadian funds to the Receiver General for Canada A deposit copy of this publication is also available for reference in public librairies across Canada Canada: $19.95 Catalog No. FS41-31/56E ISBN 0-660-10978-6 Other countries:$23.95 ISSN 0706-6481 Prices subject to change without notice Printed in Canada Thorn Press Ltd. Correct citation for this publication: THOMSON, R. E. 1981. Oceanography of the British Columbia coast. Can. Spec. Publ. Fish. Aquat. Sci. 56: 291 p. for Justine and Karen Contents FOREWORD BACKGROUND INFORMATION Introduction Acknowledgments xi Abstract/Résumé xii PART I HISTORY AND NATURE OF THE COAST Chapter 5. Upwelling: Bringing Cold Water to the Surface Chapter 1. Historical Setting Causes of Upwelling 79 Origin of the Oceans 1 Localized Effects 82 Drifting Continents 2 Climate 83 Evolution of the Coast 6 Fishing Grounds 83 Early Exploration 9 El Nifio 83 Chapter 2. -

Seymour - Belize Inlets)
Environment Canada Environnement Canada Fisheries Service des peches and Marine Service et des sciences de la mer Preliminary Catalogue of Salmon Streams and Spawning Escapements of Statistical Area 11 (Seymour - Belize Inlets) D.E. Marshal l R.F. Brown V. D. Chah ley D.G. Demontier Pac/D-77-5 Pacific Region Environment Canada Environnement Canada Fisheries Service des peches and Marine Service et des sciences de la mer Preliminary Catalogue of Salmon Streams and Spawning Escapements of Statistical Area 11 (Seymour Belize Inlets) D.E. Marshal l R.F. Brown V. D. Chahley D.G. Demontier Pac/D-77-5 Pacific Region STATISTICAL AREAS FISHERIES & MARINE SERVICE PACIFIC REGION 1110149"1"1" IIIIIIMI ' , ' I •1 ii CONTENTS Statistical Areas ii Conservation Districts iv Management Branches Salmon Spawning Streams vi Escapement Record for Statistical Area 11 vii Standards Used on Description Page viii Map References xi STREAM DATA BAMFORD CREEK, LEE CREEK & WHELAKIS CREEK 1 CHIEF NOLLIS CREEK 5 DRIFTWOOD CREEK 9 EVA CREEK 13 JAP CREEK 17 LASSITER BAY CREEK & ROWLEY BAY CREEK 21 PACK LAKE CREEK 25 QUASHELLA CREEK 29 RAINBOW CREEK 33 SCHWARTZENBERG LAGOON CREEK 37 SEYMOUR RIVER 41 TAALTZ CREEK 45 WAAMTX CREEK 49 WARNER BAY CREEK 53 WAUMP CREEK & ALISON RIVER 57 WAWATLE BAY CREEK 61 WODEFORD CREEK 65 METRIC EQUIVALENTS, WATER QUANTITIES AND FLOW MEASUREMENTS 70 ' l I CONSERVATION DISTRICTS / /FISHERIES AND MARINE SERVICE / I' / I PACIFIC REGION 1 I i 1 I ) , ) 1 — _ - r • ■•••• I 1, BRITISH COLUMBIA C. D. 10 „•'>S‘,•Li 3S ' 4't (”- ti= Cope Ih,e ,on C D.