Explanation in Biology: Reduction, Pluralism, and Explanatory Aims
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Natural Kinds and Concepts: a Pragmatist and Methodologically Naturalistic Account
Natural Kinds and Concepts: A Pragmatist and Methodologically Naturalistic Account Ingo Brigandt Abstract: In this chapter I lay out a notion of philosophical naturalism that aligns with prag- matism. It is developed and illustrated by a presentation of my views on natural kinds and my theory of concepts. Both accounts reflect a methodological naturalism and are defended not by way of metaphysical considerations, but in terms of their philosophical fruitfulness. A core theme is that the epistemic interests of scientists have to be taken into account by any natural- istic philosophy of science in general, and any account of natural kinds and scientific concepts in particular. I conclude with general methodological remarks on how to develop and defend philosophical notions without using intuitions. The central aim of this essay is to put forward a notion of naturalism that broadly aligns with pragmatism. I do so by outlining my views on natural kinds and my account of concepts, which I have defended in recent publications (Brigandt, 2009, 2010b). Philosophical accounts of both natural kinds and con- cepts are usually taken to be metaphysical endeavours, which attempt to develop a theory of the nature of natural kinds (as objectively existing entities of the world) or of the nature of concepts (as objectively existing mental entities). However, I shall argue that any account of natural kinds or concepts must an- swer to epistemological questions as well and will offer a simultaneously prag- matist and naturalistic defence of my views on natural kinds and concepts. Many philosophers conceive of naturalism as a primarily metaphysical doc- trine, such as a commitment to a physicalist ontology or the idea that humans and their intellectual and moral capacities are a part of nature. -
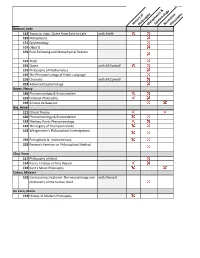
History of Philosophymetaphysics & Epistem Ology Norm Ative
HistoryPhilosophy of MetaphysicsEpistemology & NormativePhilosophy Azzouni, Jody 114 Topics in Logic: Quine from Early to Late with Smith 120 Metaphysics 131 Epistemology 191 Objects 191 Rule-Following and Metaphysical Realism 191 Truth 191 Quine with McConnell 191 Philosophy of Mathematics 192 The Phenomenology of Public Language 195 Chomsky with McConnell 292 Advanced Epistemology Bauer, Nancy 186 Phenomenology & Existentialism 191 Feminist Philosophy 192 Simone de Beauvoir Baz, Avner 121 Ethical Theory 186 Phenomenology & Existentialism 192 Merleau Ponty Phenomenology 192 The Legacy of Thompson Clarke 192 Wittgenstein's Philosophical Investigations 292 Perceptions & Indeterminacy 292 Research Seminar on Philosophical Method Choi, Yoon 117 Philosophy of Mind 164 Kant's Critique of Pure Reason 192 Kant's Moral Philosophy Cohen, Michael 192 Conciousness Explored: The neurobiology and with Dennett philosophy of the human mind De Caro, Mario 152 History of Modern Philosophy De HistoryPhilosophy of MetaphysicsEpistemology & NormativePhilosophy Car 191 Nature & Norms 191 Free Will & Moral Responsibility Denby, David 133 Philosophy of Language 152 History of Modern Philosophy 191 History of Analytic Philosophy 191 Topics in Metaphysics 195 The Philosophy of David Lewis Dennett, Dan 191 Foundations of Cognitive Science 192 Artificial Agents & Autonomy with Scheutz 192 Cultural Evolution-Is it Darwinian 192 Topics in Philosophy of Biology: The Boundaries with Forber of Darwinian Theory 192 Evolving Minds: From Bacteria to Bach 192 Conciousness -

The Myth of Supervenient Microphysicalism
The myth of supervenient microphysicalism Juan Diego Morales1 (Universidad de Cartagena) So my aim is only to establish conditional claims of the form: even if certain facts are emergent vis‑à‑vis the microphysical realm, Physicalism can still be true. Papineau, 2008: 132 1. What physicalism is Let us begin with the idea that physicalism is the ontological theory which claims that the entities that constitute our world are physical enti‑ ties, phenomena which the physical sciences must discover and articulate in their theories.2 Following the naturalistic trend of philosophy in recent times, con‑ temporary philosophers have considered that physicalism is both an a pos‑ teriori and contingent thesis. Its a posteriori sense derives from the fact of being a position that tries to overcome the problems of its direct predeces‑ 1 [email protected]. My thanks to Alejandro Rosas, Brian McLaughlin, Reinaldo Bernal, and Larry Shapiro, for helpful and clarifying comments on drafts of this paper. 2 Here I will use the general sense of “entity” and “phenomenon” for including both particulars (as objects, events, processes, and so on) and what many theorists take as universals (as properties, relations, and laws). Philosophica, 48, Lisboa, 2016, pp. 47-61. Philosophica 48.indd 47 22/11/2016 9:26:50 PM 48 Juan Diego Morales sor, materialism. The latter was established as a metaphysical doctrine that attempted to specify the entities of our world in an a priori way, in terms of a specific set of features that supposedly defined the material; features such as conservation, deterministic and on contact interaction, impenetrability, inertia, and solidity.3 But this a priori specification proved to be wrong. -

Articles Revisiting the Logical Empiricist Criticisms of Vitalism Bohang Chen1 Abstract
Revisiting the Logical Empiricist Criticisms of Vitalism Bohang Chen Transversal: International Journal for the Historiography of Science 2019 (7): 1-17 ISSN 2526-2270 www.historiographyofscience.org Belo Horizonte – MG / Brazil © The Author 2019 – This is an open access article Articles Revisiting the Logical Empiricist Criticisms of Vitalism Bohang Chen1 Abstract: Vitalism claims that biological organisms are governed by nonmaterial agents like entelechies. The received view today rejects vitalism by presupposing metaphysical materialism (or physicalism). Metaphysical materialism maintains that the world is ultimately material (or physical), and it, therefore, repudiates the existence of nonmaterial entelechies. However, this marks a shift compared with the arguments against vitalism developed by logical empiricists, who were indifferent to metaphysical issues and were only concerned with logical and empirical matters in the sciences. Logical empiricists rejected the concept of the entelechy (vitalism), because vital laws confirmed by biological phenomena were unavailable; in contrast, they accepted the concept of the atom (materialism), since it constituted physical laws and was therefore associated with verifiable results in modern physics. 1 Keywords: Vitalism; Materialism (or Physicalism); Logical Empiricism; Entelechy; Atom Received: 27 June 2019. Reviewed: 13 October 2019. Accepted: 17 October 2019. DOI: http://dx.doi.org/10.24117/2526-2270.2019.i7.03 This work is licensed under a Creative Commons Attribution 4.0 International License. _______________________________________________________________________ Introduction: The Metaphysical Refutation of Vitalism Today most biologists and philosophers understand vitalism as a heretical doctrine in the history of biology, originally proposed by Hans Driesch in the early twentieth century. 2 According to Driesch’s doctrine of vitalism, biological organisms are governed by nonmaterial vital agents termed entelechies (Driesch 1929; Churchill 1969; Allen 2008). -

Linguistics As a “Special Science”: a Comparison of Sapir and Fodor Els Elffers
Chapter 4 Linguistics as a “special science”: A comparison of Sapir and Fodor Els Elffers Independently of each other, the linguist-anthropologist Edward Sapir (1884–1939) and the philosopher of mind Jerry Fodor (1935–2017) developed a similar typol- ogy of scientific disciplines. “Basic” (Fodor) or “conceptual” (Sapir) sciences (e.g. physics) are distinguished from “special” (Fodor) or “historical” (Sapir) sciences (e.g. linguistics). Ontologically, the latter sciences are reducible to the former, but they keep their autonomy as intellectual enterprises, because their “natural kinds” are unlike those of the basic sciences. Fodor labelled this view “token physicalism”. Although Sapir’s and Fodor’s ideas were presented in very different periods of intel- lectual history (in 1917 and 1974) and in very different intellectual contexts (roughly: Geisteswissenschaften and logical positivism), the similarity between them is strik- ing. When compared in detail, some substantial differences can also be observed, which are mainly related to contextual differences. When applied to linguistics, Sapir’s and Fodor’s views offer a perspective of autonomy, albeit in different ways: for Fodor, but not for Sapir, linguistics is a subfield of psychology. 1 Introduction In 1974, Jerry Fodor (1935–2017) introduced “token physicalism”, a non-reductive variety of physicalism, which applies to “special sciences”.1 According to Fodor, special sciences, such as economics, psychology and linguistics cannot be en- tirely reduced to physics, which is a “basic science”. Such a reduction would im- ply that special sciences actually disappear as autonomous sciences. According to Fodor, special sciences retain their autonomy, because reduc- tion is possible only with respect to the events they describe (“tokens”), not with 1Token physicalism belongs to a larger class of non-reductive types of physicalism. -

On Interpreting Strong Supervenience
Western Michigan University ScholarWorks at WMU Master's Theses Graduate College 12-1996 On Interpreting Strong Supervenience Zachary J. Ernst Follow this and additional works at: https://scholarworks.wmich.edu/masters_theses Part of the Philosophy Commons Recommended Citation Ernst, Zachary J., "On Interpreting Strong Supervenience" (1996). Master's Theses. 3475. https://scholarworks.wmich.edu/masters_theses/3475 This Masters Thesis-Open Access is brought to you for free and open access by the Graduate College at ScholarWorks at WMU. It has been accepted for inclusion in Master's Theses by an authorized administrator of ScholarWorks at WMU. For more information, please contact [email protected]. ON INTERPRETING STRONG SUPERVENIENCE by Zachary J. Ernst A Thesis Submitted to the Faculty of The Graduate College in partial fulfillmentof the requirements for the Degree of Master of Arts Department of Philosophy Western Michigan University Kalamazoo, Michigan December 1996 Copyright by Zachary J. Ernst 1996 ON INTERPRETING STRONG SUPERVENIENCE Zachary J. Ernst, M.A. WesternMichigan University,_ 1996 J aegwon Kim's definitionof strong supervenience has found application in such areas as the mind-body problem, aesthetics, morality, and the relationship between physics and the special sciences. The main reason forthe popularity of supervenience is that it purportedly has a long laundry list of virtues. For instance, it has been claimed that supervenience accounts are non-reductive, capable of empirical verification, simple with respect to ontology, and explanatorily powerful. In this paper, I examine Kim's definition of strong supervenience, arguing that a fundamental ambiguity in the definition makes it impossible for strong supervenience to possess all of these virtues simultaneously. -

Philosophy of Science Reading List
Philosophy of Science Area Comprehensive Exam Reading List Revised September 2011 Exam Format: Students will have four hours to write answers to four questions, chosen from a list of approximately 20-30 questions organized according to topic: I. General Philosophy of Science II. History of Philosophy of Science III. Special Topics a. Philosophy of Physics b. Philosophy of Biology c. Philosophy of Mind / Cognitive Science d. Logic and Foundations of Mathematics Students are required to answer a total of three questions from sections I and II (at least one from each section), and one question from section III. For each section, we have provided a list of core readings—mostly journal articles and book chapters—that are representative of the material with which we expect you to be familiar. Many of these readings will already be familiar to you from your coursework and other reading. Use this as a guide to filling in areas in which you are less well- prepared. Please note, however, that these readings do not constitute necessary or sufficient background to pass the comp. The Philosophy of Science area committee assumes that anyone who plans to write this exam has a good general background in the area acquired through previous coursework and independent reading. Some anthologies There are several good anthologies of Philosophy of Science that will be useful for further background (many of the articles listed below are anthologized; references included in the list below). Richard Boyd, Philip Gasper, and J.D. Trout, eds., The Philosophy of Science (MIT Press, 991). Martin Curd and J. -

Emergence in Science and Philosophy Edited by Antonella
Emergence in Science and Philosophy Edited by Antonella Corradini and Timothy O’Connor New York London Contents List of Figures ix Introduction xi ANTONELLA CORRADINI AND TIMOTHY O’CONNOR PART I Emergence: General Perspectives Part I Introduction 3 ANTONELLA CORRADINI AND TIMOTHY O’CONNOR 1 The Secret Lives of Emergents 7 HONG YU WONG 2 On the Implications of Scientifi c Composition and Completeness: Or, The Troubles, and Troubles, of Non- Reductive Physicalism 25 CARL GILLETT 3 Weak Emergence and Context-Sensitive Reduction 46 MARK A. BEDAU 4 Two Varieties of Causal Emergentism 64 MICHELE DI FRANCESCO 5 The Emergence of Group Cognition 78 GEORG THEINER AND TIMOTHY O’CONNOR vi Contents PART II Self, Agency, and Free Will Part II Introduction 121 ANTONELLA CORRADINI AND TIMOTHY O’CONNOR 6 Why My Body is Not Me: The Unity Argument for Emergentist Self-Body Dualism 127 E. JONATHAN LOWE 7 What About the Emergence of Consciousness Deserves Puzzlement? 149 MARTINE NIDA-RÜMELIN 8 The Emergence of Rational Souls 163 UWE MEIXNER 9 Are Deliberations and Decisions Emergent, if Free? 180 ACHIM STEPHAN 10 Is Emergentism Refuted by the Neurosciences? The Case of Free Will 190 MARIO DE CARO PART III Physics, Mathematics, and the Special Sciences Part III Introduction 207 ANTONELLA CORRADINI AND TIMOTHY O’CONNOR 11 Emergence in Physics 213 PATRICK MCGIVERN AND ALEXANDER RUEGER 12 The Emergence of the Intuition of Truth in Mathematical Thought 233 SERGIO GALVAN Contents vii 13 The Emergence of Mind at the Co-Evolutive Level 251 ARTURO CARSETTI 14 Emerging Mental Phenomena: Implications for Psychological Explanation 266 ALESSANDRO ANTONIETTI 15 How Special Are Special Sciences? 289 ANTONELLA CORRADINI Contributors 305 Index 309 Introduction Antonella Corradini and Timothy O’Connor The concept of emergence has seen a signifi cant resurgence in philosophy and a number of sciences in the past couple decades. -

Conceptual Engineering and Conceptual Ethics OUP CORRECTED PROOF – FINAL, 17/12/2019, Spi OUP CORRECTED PROOF – FINAL, 17/12/2019, Spi
OUP CORRECTED PROOF – FINAL, 17/12/2019, SPi Conceptual Engineering and Conceptual Ethics OUP CORRECTED PROOF – FINAL, 17/12/2019, SPi OUP CORRECTED PROOF – FINAL, 17/12/2019, SPi Conceptual Engineering and Conceptual Ethics Alexis Burgess, Herman Cappelen, and David Plunkett 1 OUP CORRECTED PROOF – FINAL, 17/12/2019, SPi 3 Great Clarendon Street, Oxford, OX2 6DP, United Kingdom Oxford University Press is a department of the University of Oxford. It furthers the University’s objective of excellence in research, scholarship, and education by publishing worldwide. Oxford is a registered trade mark of Oxford University Press in the UK and in certain other countries © the several contributors 2020 The moral rights of the authors have been asserted First Edition published in 2020 Impression: 1 Some rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, for commercial purposes, without the prior permission in writing of Oxford University Press, or as expressly permitted by law, by licence or under terms agreed with the appropriate reprographics rights organization. This is an open access publication, available online and distributed under the terms of a Creative Commons Attribution – Non Commercial – No Derivatives 4.0 International licence (CC BY-NC-ND 4.0), a copy of which is available at http://creativecommons.org/licenses/by-nc-nd/4.0/. Enquiries concerning reproduction outside the scope of this licence should be sent to the Rights Department, Oxford University Press, at the address above Published in the United States of America by Oxford University Press 198 Madison Avenue, New York, NY 10016, United States of America British Library Cataloguing in Publication Data Data available Library of Congress Control Number: 2019946746 ISBN 978–0–19–880185–6 DOI: 10.1093/oso/9780198801856.003.0001 Printed and bound in Great Britain by Clays Ltd, Elcograf S.p.A. -

Reductionism in Biology
pdf version of the entry Reductionism in Biology http://plato.stanford.edu/archives/sum2012/entries/reduction-biology/ Reductionism in Biology from the Summer 2012 Edition of the First published Tue May 27, 2008; substantive revision Mon Apr 30, 2012 Stanford Encyclopedia Reductionism encompasses a set of ontological, epistemological, and methodological claims about the relations between different scientific of Philosophy domains. The basic question of reduction is whether the properties, concepts, explanations, or methods from one scientific domain (typically at higher levels of organization) can be deduced from or explained by the properties, concepts, explanations, or methods from another domain of science (typically one about lower levels of organization). Reduction is Edward N. Zalta Uri Nodelman Colin Allen John Perry germane to a variety of issues in philosophy of science, including the Principal Editor Senior Editor Associate Editor Faculty Sponsor structure of scientific theories, the relations between different scientific Editorial Board disciplines, the nature of explanation, the diversity of methodology, and http://plato.stanford.edu/board.html the very idea of theoretical progress, as well as to numerous topics in Library of Congress Catalog Data metaphysics and philosophy of mind, such as emergence, mereology, and ISSN: 1095-5054 supervenience. Notice: This PDF version was distributed by request to mem- In recent philosophy of biology (1970s to the 1990s), the primary debate bers of the Friends of the SEP Society and by courtesy to SEP content contributors. It is solely for their fair use. Unauthorized about reduction has focused on the question of whether and in what sense distribution is prohibited. To learn how to join the Friends of the classical genetics can be reduced to molecular biology. -

The Philosophy of Biology Edited by David L
Cambridge University Press 978-0-521-85128-2 - The Cambridge Companion to the Philosophy of Biology Edited by David L. Hull and Michael Ruse Frontmatter More information the cambridge companion to THE PHILOSOPHY OF BIOLOGY The philosophy of biology is one of the most exciting new areas in the field of philosophy and one that is attracting much attention from working scientists. This Companion, edited by two of the founders of the field, includes newly commissioned essays by senior scholars and by up-and- coming younger scholars who collectively examine the main areas of the subject – the nature of evolutionary theory, classification, teleology and function, ecology, and the prob- lematic relationship between biology and religion, among other topics. Up-to-date and comprehensive in its coverage, this unique volume will be of interest not only to professional philosophers but also to students in the humanities and researchers in the life sciences and related areas of inquiry. David L. Hull is an emeritus professor of philosophy at Northwestern University. The author of numerous books and articles on topics in systematics, evolutionary theory, philosophy of biology, and naturalized epistemology, he is a recipient of a Guggenheim Foundation fellowship and is a Fellow of the American Academy of Arts and Sciences. Michael Ruse is professor of philosophy at Florida State University. He is the author of many books on evolutionary biology, including Can a Darwinian Be a Christian? and Darwinism and Its Discontents, both published by Cam- bridge University Press. A Fellow of the Royal Society of Canada and the American Association for the Advancement of Science, he has appeared on television and radio, and he contributes regularly to popular media such as the New York Times, the Washington Post, and Playboy magazine. -

May 03 CI.Qxd
Philosophy of Chemistry by Eric Scerri Of course, the field of compu- hen I push my hand down onto my desk it tational quantum chemistry, does not generally pass through the which Dirac and other pioneers Wwooden surface. Why not? From the per- of quantum mechanics inadver- spective of physics perhaps it ought to, since we are tently started, has been increas- told that the atoms that make up all materials consist ingly fruitful in modern mostly of empty space. Here is a similar question that chemistry. But this activity has is put in a more sophisticated fashion. In modern certainly not replaced the kind of physics any body is described as a superposition of chemistry in which most practitioners are engaged. many wavefunctions, all of which stretch out to infin- Indeed, chemists far outnumber scientists in all other ity in principle. How is it then that the person I am fields of science. According to some indicators, such as talking to across my desk appears to be located in one the numbers of articles published per year, chemistry particular place? may even outnumber all the other fields of science put together. If there is any sense in which chemistry can The general answer to both such questions is that be said to have been reduced then it can equally be although the physics of microscopic objects essen- said to be in a high state of "oxidation" regarding cur- tially governs all of matter, one must also appeal to rent academic and industrial productivity. the laws of chemistry, material science, biology, and Starting in the early 1990s a group of philosophers other sciences.