The Effect of Trait Anxiety on the Generalisation of Fear Acquisition and Extinction
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Greg J Sheehan, FWS Principal Deputy Director
Greg J Sheehan, FWS Principal Deputy Director Tue Jun 6, 2017 2:45pm FWS weekly check in meeting Video call: (b) (5), (b) (6) Where: AS/FWP Conference Room -- 3144 Calendar: Maureen Foster Created by: Tasha Robbins Who: Thomas Irwin, Charisa Morris, Maureen Foster, Richard Goeken, GregSheehan, Wendy Fink, Stephen Guertin, Catherine Gulac, Jason Larrabee,Tasha Robbins, Aurelia Skipwith, Marshall Critchfield, Heather Swift,Jim Kurth, Todd Willens, Roslyn Sellars, Zachariah Gambill, PegRomanik, Barbara Wainman 3pm *FWS weekly check in meeting Video call: (b) (5), (b) (6) Where: AS/FWP Conference Room -- 3144 Calendar: Greg Sheehan Created by: Roslyn Sellars Wed Jun 7, 2017 1pm OIG monthly status update meeting/conference call (Jim, Steve, Kathy Garrity, Keith Toomey, Charisa) Room 3357 Video call: (b) (5), (b) (6) Where: Dial: (b) (5) , Code: (b) (5) Calendar: Jim Kurth Created by: Roslyn Sellars Who: Casey Hammond, Stephen Guertin, Katherine Garrity, Jim Kurth, GregSheehan, Charisa Morris, Keith Toomey 2:30pm [Asst Directors/Regional Directors Only-No Deputies or Actings] - Weekly Directorate VTC: Transition Check-In--Room 3038 Video call: (b) (5), (b) (6) Calendar: Jim Kurth Created by: Thomas Irwin Who: [email protected], Tom Melius, [email protected], WandaCantrell, Edward Grace, [email protected], Henry Schlitzer,Pamela Michalegko, Kenneth Taylor, Benjamin Tuggle, Brian Bloodsworth,Robyn Thorson, Gary Frazer, Michael Gale, Jim Kurth, Charisa Morris,Paul Rauch, Denise Thompson, Seth Mott, Cynthia Martinez,cynthia_dohner@fws -
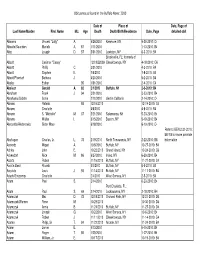
Obituaries Buffalo News 2010 by Name
Obituaries as found in the Buffalo News: 2010 Date of Place of Date, Page of Last Name/Maiden First Name M.I. Age Death Death/Birth/Residence Date, Page detailed obit Abbarno Vincent "Lolly" A. 9/26/2010 Kenmore, NY 9-30-2010: C4 Abbatte/Saunders Murielle A. 87 1/11/2010 1-13-2010: B4 Abbo Joseph D. 57 5/31/2010 Lewiston, NY 6-3-2010: B4 Brooksville, FL; formerly of Abbott Casimer "Casey" 12/19/22009 Cheektowaga, NY 4-18-2010: C6 Abbott Phillip C. 3/31/2010 4-3-2010: B4 Abbott Stephen E. 7/6/2010 7-8-2010: B4 Abbott/Pfoetsch Barbara J. 4/20/2010 5-2-2010: B4 Abeles Esther 95 1/31/2010 2-4-2010: C4 Abelson Gerald A. 82 2/1/2010 Buffalo, NY 2-3-2010: B4 Abraham Frank J. 94 3/21/2010 3-23-2010: B4 Abrahams/Gichtin Sonia 2/10/2010 died in California 2-14-2010: C4 Abramo Rafeala 93 12/16/2010 12-19-2010: C4 Abrams Charlotte 4/6/2010 4-8-2010: B4 Abrams S. "Michelle" M. 37 5/21/2010 Salamanca, NY 5-23-2010: B4 Abrams Walter I. 5/15/2010 Basom, NY 5-19-2010: B4 Abrosette/Aksterowicz Sister Mary 6/18/2010 6-19-2010: C4 Refer to BEN 2-21-2010: B6/7/8 for more possible Abshagen Charles, Jr. L. 73 2/19/2010 North Tonawanda, NY 2-22-2010: B8 information Acevedo Miguel A. 10/6/2010 Buffalo, NY 10-27-2010: B4 Achkar John E. -

Vol. 83 Wednesday, No. 211 October 31, 2018 Pages 54663–54860
Vol. 83 Wednesday, No. 211 October 31, 2018 Pages 54663–54860 OFFICE OF THE FEDERAL REGISTER VerDate Sep 11 2014 20:07 Oct 30, 2018 Jkt 247001 PO 00000 Frm 00001 Fmt 4710 Sfmt 4710 E:\FR\FM\31OCWS.LOC 31OCWS amozie on DSK3GDR082PROD with FRONT MATTER WS II Federal Register / Vol. 83, No. 211 / Wednesday, October 31, 2018 The FEDERAL REGISTER (ISSN 0097–6326) is published daily, SUBSCRIPTIONS AND COPIES Monday through Friday, except official holidays, by the Office PUBLIC of the Federal Register, National Archives and Records Administration, under the Federal Register Act (44 U.S.C. Ch. 15) Subscriptions: and the regulations of the Administrative Committee of the Federal Paper or fiche 202–512–1800 Register (1 CFR Ch. I). The Superintendent of Documents, U.S. Assistance with public subscriptions 202–512–1806 Government Publishing Office, is the exclusive distributor of the official edition. Periodicals postage is paid at Washington, DC. General online information 202–512–1530; 1–888–293–6498 Single copies/back copies: The FEDERAL REGISTER provides a uniform system for making available to the public regulations and legal notices issued by Paper or fiche 202–512–1800 Federal agencies. These include Presidential proclamations and Assistance with public single copies 1–866–512–1800 Executive Orders, Federal agency documents having general (Toll-Free) applicability and legal effect, documents required to be published FEDERAL AGENCIES by act of Congress, and other Federal agency documents of public Subscriptions: interest. Assistance with Federal agency subscriptions: Documents are on file for public inspection in the Office of the Federal Register the day before they are published, unless the Email [email protected] issuing agency requests earlier filing. -

Magazin Wirtschaft
www.stuttgart.ihk.de 07.2013 Stuttgart - Böblingen - Esslingen-Nürtingen - Göppingen - Ludwigsburg - Rems-Murr Magazin Wirtschaft Ein Service der IHK für Unternehmen in der Region Stuttgart Fachkräfte finden zwischen Alb und Gäu Seite 6 wer hat schon ein ganzes kabinett nur für diewirtschaft? Selbstverständlich wir. Damit die Betriebe gute Rahmenbedingungen haben, sind in unseren Gremien ausschließlich Wirtschaftexperten vertreten. www.stuttgart.ihk.de oder Infoline 0711 2005-0 EDITORIAL Handelskriege kennen keine Gewinner Ähnlich dem Frühjahrswetter in diesem weit kommen? China ist die zweitgrößte Wirt- Jahr ziehen nun auch Wolken über dem schaftsmacht der Welt. Daher sollte das Land Parkett des internationalen Handels beginnen, sich an internationale Spielregeln auf. Auslöser ist die vorläufige Erhebung von zu halten. Globalisierung benötigt faire Wett- EU-Strafzöllen auf chinesische Solarmodule. bewerbsbedingungen, an die sich China zu- China kontert mit einem Anti-Dumping-Ver- weilen jedoch nicht hält. Belegte Fälle von fahren gegen europäischen Wein. Immer Wettbewerbsverstößen gibt es genug. mehr Produkte treten in den Fokus der Dum- ping-Diskussion. Vernünftige Handelspolitik Die EU muss schnellstens sieht anders aus. Mangels Deeskalation wird Verhandlungen aufnehmen die Situation für China und Europa immer tassilo zywietz unangenehmer. Ein Handelskrieg droht. Europa darf davor die Augen nicht ver- Geschäftsführer International Globalisierung erfordert jedoch gegensei- schließen. Nachgiebigkeit ist keine Erfolg ver- der IHK Region Stuttgart tige Rücksichtnahme und Handeln im Sinne sprechende Strategie – damit würde die EU der immer stärker miteinander verbundenen erpressbar. Lässt man China gewähren, kann Volkswirtschaften: China und die EU haben die Methode künftig in der Chemiebranche letztes Jahr Waren und Dienstleistungen in und irgendwann im Autobau angewendet Höhe von über 400 Milliarden Euro ausge- werden. -

2013 Kaiser Permanente-Authored Publications Alphabetical by Author
2013 Kaiser Permanente-Authored Publications Alphabetical by Author 1. Abbas MA, Cannom RR, Chiu VY, Burchette RJ, Radner GW, Haigh PI, Etzioni DA Triage of patients with acute diverticulitis: are some inpatients candidates for outpatient treatment? Colorectal Dis. 2013 Apr;15(4):451-7. Southern California 23061533 KP author(s): Abbas, Maher A; Chiu, Vicki Y; Burchette, Raoul J; Radner, Gary W; Haigh, Philip I 2. Abbass MA, Slezak JM, DiFronzo LA Predictors of Early Postoperative Outcomes in 375 Consecutive Hepatectomies: A Single- institution Experience Am Surg. 2013 Oct;79(10):961-7. Southern California 24160779 KP author(s): Abbass, Mohammad Ali; Slezak, Jeffrey M; DiFronzo, Andrew L 3. Abbenhardt C, Poole EM, Kulmacz RJ, Xiao L, Curtin K, Galbraith RL, Duggan D, Hsu L, Makar KW, Caan BJ, Koepl L, Owen RW, Scherer D, Carlson CS, Genetics and Epidemiology of Colorectal Cancer Consortium (GECCO) and CCFR, Potter JD, Slattery ML, Ulrich CM Phospholipase A2G1B polymorphisms and risk of colorectal neoplasia Int J Mol Epidemiol Genet. 2013 Sep 12;4(3):140-9. Northern California 24046806 KP author(s): Caan, Bette 4. Aberg JA, Gallant JE, Ghanem KG, Emmanuel P, Zingman BS, Horberg MA Primary Care Guidelines for the Management of Persons Infected With HIV: 2013 Update by the HIV Medicine Association of the Infectious Diseases Society of America Clin Infect Dis. 2013 Nov 13. Mid-Atlantic 24235263 KP author(s): Horberg, Michael 5. Abraham AG, D'Souza G, Jing Y, Gange SJ, Sterling TR, Silverberg MJ, Saag MS, Rourke SB, Rachlis A, Napravnik S, Moore RD, Klein MB, Kitahata MM, Kirk GD, Hogg RS, Hessol NA, Goedert JJ, Gill MJ, Gebo KA, Eron JJ, Engels EA, Dubrow R, Crane HM, Brooks JT, Bosch RJ, Strickler HD, North American AIDS Cohort Collaboration on Research and Design of IeDEA Invasive cervical cancer risk among HIV-infected women: A North American multi-cohort collaboration prospective study J Acquir Immune Defic Syndr. -

Elecciones Anuales Grado 2020 − Delegados De Curso Página 1 De 51
Elecciones anuales Grado 2020 − Delegados de curso Fecha de impresión: 27−11−2020 Grado en Ingeniería Eléctrica − Curso: 1 Nº Apellido1 Apellido2 Nombre DNI 1 ARIAS RODRIGUEZ GABRIEL 7***03*3 2 CAMPOS LUCAS ALEJANDRO 7*18***9 3 FERNANDEZ MIERA ADRIAN *2***144 4 GARCÍA GARCÍA IÑAKI 20*0*3** 5 GARCIA HERRANZ JULIO *2*94**8 6 GONZALEZ VENERO HERNAN **08*08* 7 GUERRA SAINZ FRANCISCO RAMON **104**6 8 MADARIAGA GONZALEZ ERNESTO 7*103*** 9 MARCANO CUBILLAS MANUEL 7**2*0*1 10 MAZO LOPEZ PEDRO PABLO DEL **0*26*0 11 MOYA MANTECON LAURA 7*1**15* 12 ORTIZ SAMPERIO VICTOR *21***11 13 PEREDA SOLARES JORGE **1*501* 14 RAMOS MARTINEZ SERGIO 7**8**77 15 RODRIGUEZ EIZAGUIRRE JORGE 7*0**1*8 16 ROJAS LOPEZ FELIPE ***00*03 17 SOLANO RUIZ ALVARO ***0319* 18 XIANG QIXIN ****6291 En cumplimiento de lo dispuesto apartado 1 de la Disposición adicional séptima de la Ley Orgánica 3/2018, de 5 de diciembre, de Protección de Datos Personales y garantía de los derechos digitales, las personas que figuran en este listado aparecen identificadas con cuatro cifras numéricas aleatorias del DNI, NIE, Pasaporte o documento equivalente. En los casos en que el número de estos documentos tenga menos de seis cifras numéricas, se asignará un número aleatorio confeccionado con este fin. Los números aleatorios asignados pueden consultarse en Campus Virtual (Menú general / Datos personales). Página 1 de 51 Elecciones anuales Grado 2020 − Delegados de curso Fecha de impresión: 27−11−2020 Grado en Ingeniería Eléctrica − Curso: 2 Nº Apellido1 Apellido2 Nombre DNI 1 ALONSO MURGA RAUL -

Family Group Sheets Surname Index
PASSAIC COUNTY HISTORICAL SOCIETY FAMILY GROUP SHEETS SURNAME INDEX This collection of 660 folders contains over 50,000 family group sheets of families that resided in Passaic and Bergen Counties. These sheets were prepared by volunteers using the Societies various collections of church, ceme tery and bible records as well as city directo ries, county history books, newspaper abstracts and the Mattie Bowman manuscript collection. Example of a typical Family Group Sheet from the collection. PASSAIC COUNTY HISTORICAL SOCIETY FAMILY GROUP SHEETS — SURNAME INDEX A Aldous Anderson Arndt Aartse Aldrich Anderton Arnot Abbott Alenson Andolina Aronsohn Abeel Alesbrook Andreasen Arquhart Abel Alesso Andrews Arrayo Aber Alexander Andriesse (see Anderson) Arrowsmith Abers Alexandra Andruss Arthur Abildgaard Alfano Angell Arthurs Abraham Alje (see Alyea) Anger Aruesman Abrams Aljea (see Alyea) Angland Asbell Abrash Alji (see Alyea) Angle Ash Ack Allabough Anglehart Ashbee Acker Allee Anglin Ashbey Ackerman Allen Angotti Ashe Ackerson Allenan Angus Ashfield Ackert Aller Annan Ashley Acton Allerman Anners Ashman Adair Allibone Anness Ashton Adams Alliegro Annin Ashworth Adamson Allington Anson Asper Adcroft Alliot Anthony Aspinwall Addy Allison Anton Astin Adelman Allman Antoniou Astley Adolf Allmen Apel Astwood Adrian Allyton Appel Atchison Aesben Almgren Apple Ateroft Agar Almond Applebee Atha Ager Alois Applegate Atherly Agnew Alpart Appleton Atherson Ahnert Alper Apsley Atherton Aiken Alsheimer Arbuthnot Atkins Aikman Alterman Archbold Atkinson Aimone -

Electrophysiology Read-Out Tools for Brain-On-Chip Biotechnology
micromachines Review Electrophysiology Read-Out Tools for Brain-on-Chip Biotechnology Csaba Forro 1,2,†, Davide Caron 3,† , Gian Nicola Angotzi 4,†, Vincenzo Gallo 3, Luca Berdondini 4 , Francesca Santoro 1 , Gemma Palazzolo 3,* and Gabriella Panuccio 3,* 1 Tissue Electronics, Fondazione Istituto Italiano di Tecnologia, Largo Barsanti e Matteucci, 53-80125 Naples, Italy; [email protected] (C.F.); [email protected] (F.S.) 2 Department of Chemistry, Stanford University, Stanford, CA 94305, USA 3 Enhanced Regenerative Medicine, Fondazione Istituto Italiano di Tecnologia, Via Morego, 30-16163 Genova, Italy; [email protected] (D.C.); [email protected] (V.G.) 4 Microtechnology for Neuroelectronics, Fondazione Istituto Italiano di Tecnologia, Via Morego, 30-16163 Genova, Italy; [email protected] (G.N.A.); [email protected] (L.B.) * Correspondence: [email protected] (G.P.); [email protected] (G.P.); Tel.: +39-010-2896-884 (G.P.); +39-010-2896-493 (G.P.) † These authors contributed equally to this paper. Abstract: Brain-on-Chip (BoC) biotechnology is emerging as a promising tool for biomedical and pharmaceutical research applied to the neurosciences. At the convergence between lab-on-chip and cell biology, BoC couples in vitro three-dimensional brain-like systems to an engineered microfluidics platform designed to provide an in vivo-like extrinsic microenvironment with the aim of replicating tissue- or organ-level physiological functions. BoC therefore offers the advantage of an in vitro repro- duction of brain structures that is more faithful to the native correlate than what is obtained with conventional cell culture techniques. -

C9orf72 Frontotemporal Dementia and Amyotrophic Lateral Sclerosis: Investigating Repeat Pathology in Cell Culture Models and Human Post-Mortem Brain
C9orf72 frontotemporal dementia and amyotrophic lateral sclerosis: investigating repeat pathology in cell culture models and human post-mortem brain. Charlotte Elizabeth Ridler A thesis submitted in partial fulfilment of the requirements for the degree of Doctor of Philosophy from University College London Department of Neurodegenerative Disease Institute of Neurology University College London 2016 1 Declaration I, Charlotte Ridler, confirm that the work presented in this thesis is my own. Where information has been derived from other sources, I confirm that this has been indicated in the thesis. 2 Acknowledgements My deepest thanks go to everyone who has helped and supported me during this PhD project, without whom it would not have been possible. Thank you firstly to my supervisor Dr. Adrian Isaacs for all of his guidance, encouragement and enthusiastic discussions during this exciting and unpredictable project. Thank you also to Dr. Sarah Mizielinska, who has been an incredible mentor and role-model during this PhD; I have learnt a huge amount from her endless dedication and generosity. I would also like to thank my secondary supervisor Prof. Elizabeth Fisher for all her wise words of advice. Thank you to all members of the Isaacs and Fisher labs past and present. Special thanks to Frances Norona, Dr. Karen Cleverly, Julian Pietrzyk, Dr. Ione Woollacott and Dr. Anny Devoy who were all great help during the cloning phase of this project; to Dr. Rubika Balendra for all her help and discussions in the nucleoli work; and to Dr. Emma Clayton and Dr. Roberto Simone for their constant support. A number of other people have lent me their valuable expertise for this project, for which I am very grateful. -

1¾ « Lõ Ô C:\WORD5\HPLASER
................................................................ INN15 IIIIIIIIIIII NNNN NN NNNN NN 11 555555 II NN NN NN NN NN NN 111 55 II NN NN NN NN NN NN 11 55 II NN NN NN NN NN NN = 11 55555 II NN NN NN NN NN NN = 11 55 II NN NN NN NN NN NN 11 55 55 IIIIIIIIIII NN NNNN NN NNNN 111111 555555 INTERNATIONAL NETWORK NEWSLETTER NUMBER 15 JUNE 1991 ============================================================ ABSTRACT Thanks are expressed to the Gang of Three who so successfully organized the Network's conference in Normal in May....announcement is made of the next Network Graduate Workshop and the next Network International Conference in Milwaukee in 1993 the recipients of the Network's initiative to support graduate student travel are recognized....the procedures for the next Network Dissertation Award are announced....The Committee on the Future of the Network reports on its proposals, along with progress report on the talks on the proposed merger with ISSPR .. a new feature of the Newsletters is announced .... a copy of the program for the Normal conference is included in case those who missed the conference would like to benefit by contacting authors of papers MAJOR ANNOUNCEMENT: IT IS TIME TO RENEW YOUR MEMBERSHIP. NOTE THAT YOU CAN GET A DISCOUNT BY RENEWING BEFORE SEPTEMBER 30TH THE "NORMAL" CONFERENCE The Normal conference was, from start to finish, a thoroughly well-organized, thoroughly stimulating and thoroughly enjoyable affair with a wonderful atmosphere free from all rancor and strife! Sound too good to be true? Well, if you were not there then ask someone who was and if you were, you probably have already told others. -

HPC Passes Resolution by BRIDGET O’CONNOR Members in Their Discussion of the Reso News Writer Lution Was the Regulation of Free Speech As It Relates to All Students
iOBSERVER Wednesday, October 30, 1996 • Vol. XXX No. 42 THE INDEPENDENT NEWSPAPER SERVING NOTRE DAME AND SAINT MARY'S m HALL PRESIDENTS' COUNCIL HPC passes resolution By BRIDGET O’CONNOR members in their discussion of the reso News Writer lution was the regulation of free speech as it relates to all students. “Everyone After the second closed door discussion pretty much agreed that the University in two consecutive meetings, the Hall should not be allowed to regulate the free Presidents’ Council voted last night to speech of students.” pass the resolution introduced by the Because the resolution contained two College Democrats last week by a simple clauses, one addressing the actual denial majority vote. Thirteen members voted in of the College Democrats’ registration to favor of the resolution, 10 voted against demonstrate and the second specifically and two abstained. aimed at the treatment of gay, lesbian Conducted using secret ballots, the vote and bisexual students’ rights, some of the was the culmination of a week of discus HPC members were unsure what a vote sion among students and between the for the resolution would signify. Some hall presidents and dorm residents. The members felt very strongly that gay and issue had been tabled at the end of the lesbian students should be specifically last HPC meeting so that the members identified in the resolution while others could discuss the issue and become more felt that it should be broadened to include familiar with the details of the incident. a call for the protection of all students’ The Observer/Michelle Sweet The HPC debated and passed a resolution presented to the council by the College According to Mike Tobin, co-president Democrats concerning student treatment by the Office of Student Affairs. -

University Staff University Staff
www.ualberta.ca UNIVERSITY OF ALBERTA University Staff University University Staff In Memoriam: Agrawal, Babita, BSc MSc (Allahabad), PhD Anderson, Colin C, BSc (Honours) PhD (Western Al-Hussaini, Ata N, BSc (Iraq), MSc (Purdue), PhD (Illinois), Professor Emeritus of Mathematical Sciences (Alberta), Assistant Professor of Surgery (2001, Ontario), Assistant Professor of Surgery (2002, (1968, 1999). 2001). 2002). Bennett, Bonnie L, Manager, Admin Services and Information Systems, Human Resources (1998, 1998). Aguerrevere, Felipe L, BS (Simon Bolivar), Anderson, Todd W, Director of the Bookstore Blackley, Frank D, MA PhD (Toronto), Professor Emeritus of History (1950, 1983). MAdmin (IESA), PhD (Calif-LA), Assistant (1999, 2001). Chamberlin, Charles R, BA (Iowa), MA (Minnesota), PhD (Minnesota), Professor Emeritus of Elementary Professor of Finance & Management Science Anderson, Wayne, Administrative Professional Education (1969, 1995). (2000, 2000). Officer in the Bookstore (1999, 1999). Faulds, Alexander G, MCSP DipTP (London), Professor Emeritus of Physical Therapy (1958, 1977). Ahmed, Syed N, MBBS (Dow Medical), Assistant Andrew, Susan E, BSc (Toronto), MSc (Simon Ferrate, Juan, Lic (Barcelona), Professor Emeritus of Comparative Literature (1962, 1985). Professor of Medicine (2002, 2002). Fraser), PhD (British Columbia), Assistant Gifford, David J, BSc PhD (Alberta), Professor of Biological Sciences (1986, 1998). Albrecht, G Diane, BComm (Alberta), Director of Professor of Medical Genetics (1998, 1998). Gilbert, James AL, MB ChB MD FRCP (Edinburgh), FRCP(C), FRCP (London), FACP, Professor Emeritus of Staff Programs in Human Resources (2000, Andrews, Debra, BSC (Wilkes), MD (Harvard), Medicine (1950, 1983). 2001). FRCP, Associate Professor of Pediatrics (1987, Gunning, Harry E, BA MA PhD (Toronto), DSc (Guelph), DSc (Queen’s), FRSC, Professor Emeritus of Chemistry Alexander, Scott DB, BSc MSc PhD (Alberta), 2003).