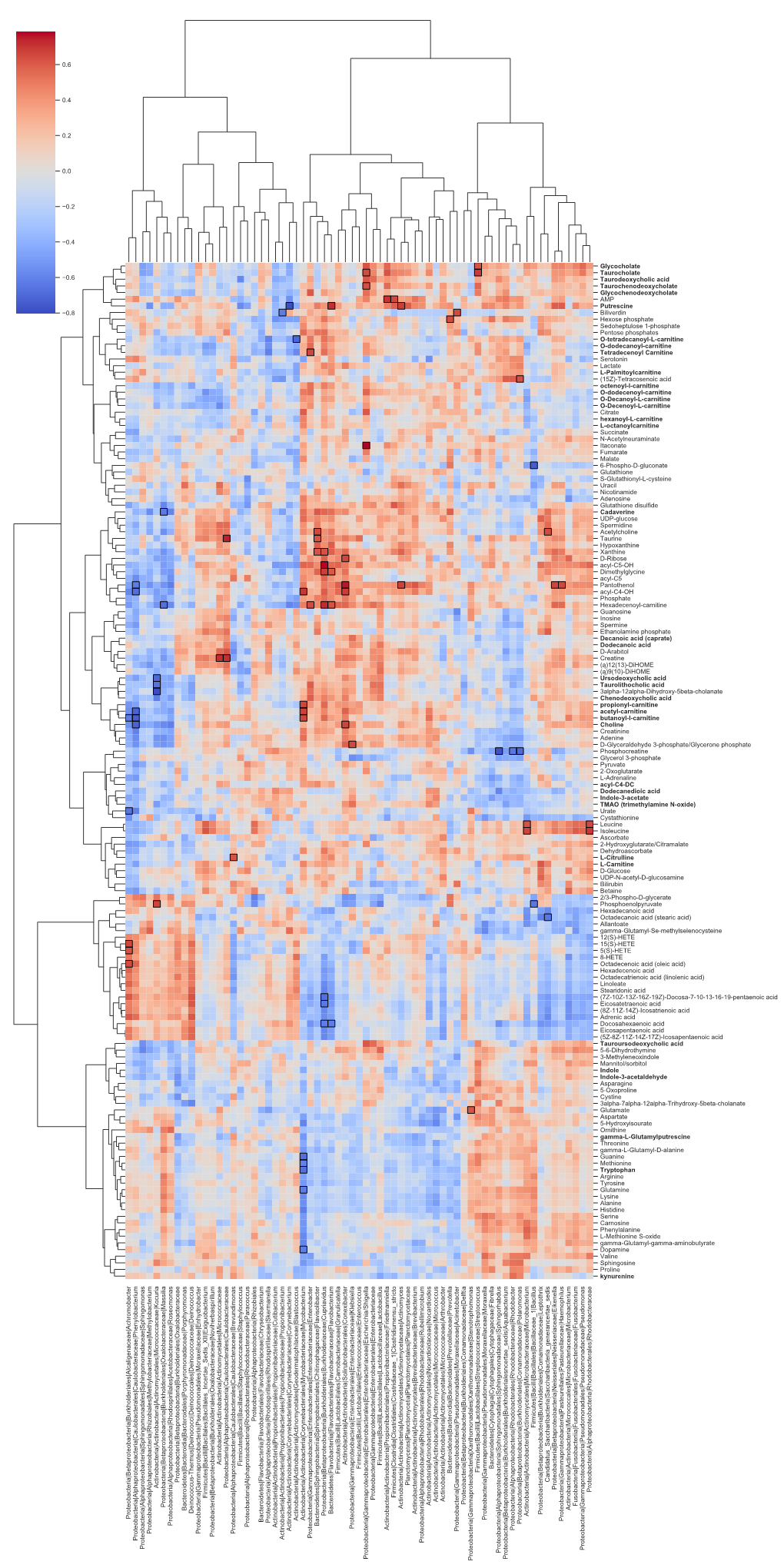
Glycocholate Taurocholate Taurodeoxycholic Acid
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Isbn 978-625-409-353-1
ISBN 978-625-409-353-1 INTERNATIONAL CONGRESS ON BIOLOGICAL AND HEALTH SCIENCES PROCEEDİNGS BOOK This work is subject to copyright and all rights reserved, whether the whole or part of the material is concerned. The right to publish this book belongs to International Congress on Biological and Health Sciences-2021. No part of this publication may be translated, reproduced, stored in a computerized system, or transmitted in any form or by any means, including, but not limited to electronic, mechanical, photocopying, recording without written permission from the publisher. This Proceedings Book has been published as an electronic publication (e-book). The publisher is not responsible for possible damages, which may be a result of content derived from this electronic publication All authors are responsible for the contents of their abstracts. https://www.biohealthcongress.com/ ([email protected]) Editor Ulaş ACARÖZ Published: 28/03/2021 ISBN: Editor's Note The first ‘International Congress on Biological and Health Sciences’ was organized online and free of charge. We are very happy and proud that various health science-related fields attended the congress. By this event, the distinguished and respected scientists came together to exchange ideas, develop and implement new researches and joint projects. There were 15 invited speakers from 10 different countries and also approximately 400 submissions were accepted from more than 20 countries. We would like to thank all participants and supporters. Hope to see you at our next congress. -

Succession and Persistence of Microbial Communities and Antimicrobial Resistance Genes Associated with International Space Stati
Singh et al. Microbiome (2018) 6:204 https://doi.org/10.1186/s40168-018-0585-2 RESEARCH Open Access Succession and persistence of microbial communities and antimicrobial resistance genes associated with International Space Station environmental surfaces Nitin Kumar Singh1, Jason M. Wood1, Fathi Karouia2,3 and Kasthuri Venkateswaran1* Abstract Background: The International Space Station (ISS) is an ideal test bed for studying the effects of microbial persistence and succession on a closed system during long space flight. Culture-based analyses, targeted gene-based amplicon sequencing (bacteriome, mycobiome, and resistome), and shotgun metagenomics approaches have previously been performed on ISS environmental sample sets using whole genome amplification (WGA). However, this is the first study reporting on the metagenomes sampled from ISS environmental surfaces without the use of WGA. Metagenome sequences generated from eight defined ISS environmental locations in three consecutive flights were analyzed to assess the succession and persistence of microbial communities, their antimicrobial resistance (AMR) profiles, and virulence properties. Metagenomic sequences were produced from the samples treated with propidium monoazide (PMA) to measure intact microorganisms. Results: The intact microbial communities detected in Flight 1 and Flight 2 samples were significantly more similar to each other than to Flight 3 samples. Among 318 microbial species detected, 46 species constituting 18 genera were common in all flight samples. Risk group or biosafety level 2 microorganisms that persisted among all three flights were Acinetobacter baumannii, Haemophilus influenzae, Klebsiella pneumoniae, Salmonella enterica, Shigella sonnei, Staphylococcus aureus, Yersinia frederiksenii,andAspergillus lentulus.EventhoughRhodotorula and Pantoea dominated the ISS microbiome, Pantoea exhibited succession and persistence. K. pneumoniae persisted in one location (US Node 1) of all three flights and might have spread to six out of the eight locations sampled on Flight 3. -
Annotation-1 Annotation-1
Annotation-1 Baseline Resuscitation Normal Saline Resuscitation PFP Shock Annotation-1 Aminoacids Arginine and proline metabolism Carnitine and fatty acid metabolsim Glutamate metabolism Glycerophospholipid biosynthesis Glycolysis and sugars GSH homeostasis GSH homeostasis/Glyoxlate Hexosamine Indole and Tryptophan Nucleotides Other Panthothenate metabolism Pentose Phosphate Pathway Serine biosynthesis and one-carbon metabolism Signaling Sulfur metabolism TCA cycle urea cycle relative row min row max Baseline_14 Baseline_16 Baseline_13 Baseline_15 Baseline_22 Baseline_2 Baseline_12 Baseline_3 Baseline_4 Baseline_9 Baseline_7 Baseline_8 Shock_13 Shock_12 Shock_15 Shock_22 Shock_14 Shock_16 Shock_2 Shock_3 Shock_7 Shock_4 Shock_8 Shock_9 Res_NS_14 Res_NS_13 Res_NS_16 Res_NS_12 Res_NS_22 Res_NS_15 Res_PFP_2 Res_PFP_3 Res_PFP_7 Res_PFP_4 Res_PFP_8 Res_PFP_9 Annotation-1 Annotation-1 Annotation Annotation-1 L-Arginine Aminoacids L-Isoleucine Aminoacids Leucine Aminoacids L-Cysteine Aminoacids L-Alanine Aminoacids L-Aspartate Aminoacids L-Glutamate Aminoacids L-Glutamine Aminoacids L-Histidine Aminoacids L-Lysine Aminoacids L-Methionine Aminoacids L-Tyrosine Aminoacids L-Asparagine Aminoacids L-Threonine Aminoacids L-Cystine Aminoacids L-Serine Aminoacids L-Proline Aminoacids L-Valine Aminoacids L-Tryptophan Aminoacids Glycine Aminoacids L-Kynurenine Aminoacids L-Phenylalanine Aminoacids CMP Nucleotides 6-Hydroxynicotinate Nucleotides 5-6-Dihydrouracil Nucleotides AMP Nucleotides dAMP Nucleotides GMP Nucleotides Guanine Nucleotides 2-5-Dihydroxypyridine -

Multilayered Horizontal Operon Transfers from Bacteria Reconstruct a Thiamine Salvage Pathway in Yeasts
Multilayered horizontal operon transfers from bacteria reconstruct a thiamine salvage pathway in yeasts Carla Gonçalvesa and Paula Gonçalvesa,1 aApplied Molecular Biosciences Unit-UCIBIO, Departamento de Ciências da Vida, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, 2829-516 Caparica, Portugal Edited by Edward F. DeLong, University of Hawaii at Manoa, Honolulu, HI, and approved September 22, 2019 (received for review June 14, 2019) Horizontal acquisition of bacterial genes is presently recognized as nisms presumed to have facilitated a transition from bacterial an important contribution to the adaptation and evolution of operon transcription to eukaryotic-style gene expression were eukaryotic genomes. However, the mechanisms underlying ex- proposed, such as gene fusion giving rise to multifunctional pro- pression and consequent selection and fixation of the prokaryotic teins (6, 23, 24), increase in intergenic distances between genes to genes in the new eukaryotic setting are largely unknown. Here we generate room for eukaryotic promoters, and independent tran- show that genes composing the pathway for the synthesis of the scription producing mRNAs with poly(A) tails have been dem- essential vitamin B1 (thiamine) were lost in an ancestor of a yeast onstrated (22). In the best documented study, which concerns a lineage, the Wickerhamiella/Starmerella (W/S) clade, known to bacterial siderophore biosynthesis operon acquired by yeasts be- harbor an unusually large number of genes of alien origin. The longing to the Wickerhamiella/Starmerella (W/S) clade, the bacte- thiamine pathway was subsequently reassembled, at least twice, rial genes acquired as an operon were shown to be functional (22). by multiple HGT events from different bacterial donors involving Thiamine, commonly known as vitamin B1, is essential for all both single genes and entire operons. -

(Batch Learning Self-Organizing Maps), to the Microbiome Analysis of Ticks
Title A novel approach, based on BLSOMs (Batch Learning Self-Organizing Maps), to the microbiome analysis of ticks Nakao, Ryo; Abe, Takashi; Nijhof, Ard M; Yamamoto, Seigo; Jongejan, Frans; Ikemura, Toshimichi; Sugimoto, Author(s) Chihiro The ISME Journal, 7(5), 1003-1015 Citation https://doi.org/10.1038/ismej.2012.171 Issue Date 2013-03 Doc URL http://hdl.handle.net/2115/53167 Type article (author version) File Information ISME_Nakao.pdf Instructions for use Hokkaido University Collection of Scholarly and Academic Papers : HUSCAP A novel approach, based on BLSOMs (Batch Learning Self-Organizing Maps), to the microbiome analysis of ticks Ryo Nakao1,a, Takashi Abe2,3,a, Ard M. Nijhof4, Seigo Yamamoto5, Frans Jongejan6,7, Toshimichi Ikemura2, Chihiro Sugimoto1 1Division of Collaboration and Education, Research Center for Zoonosis Control, Hokkaido University, Kita-20, Nishi-10, Kita-ku, Sapporo, Hokkaido 001-0020, Japan 2Nagahama Institute of Bio-Science and Technology, Nagahama, Shiga 526-0829, Japan 3Graduate School of Science & Technology, Niigata University, 8050, Igarashi 2-no-cho, Nishi- ku, Niigata 950-2181, Japan 4Institute for Parasitology and Tropical Veterinary Medicine, Freie Universität Berlin, Königsweg 67, 14163 Berlin, Germany 5Miyazaki Prefectural Institute for Public Health and Environment, 2-3-2 Gakuen Kibanadai Nishi, Miyazaki 889-2155, Japan 6Utrecht Centre for Tick-borne Diseases (UCTD), Department of Infectious Diseases and Immunology, Faculty of Veterinary Medicine, Utrecht University, Yalelaan 1, 3584 CL Utrecht, The Netherlands 7Department of Veterinary Tropical Diseases, Faculty of Veterinary Science, University of Pretoria, Private Bag X04, 0110 Onderstepoort, South Africa aThese authors contributed equally to this work. Keywords: BLSOMs/emerging diseases/metagenomics/microbiomes/symbionts/ticks Running title: Tick microbiomes revealed by BLSOMs Subject category: Microbe-microbe and microbe-host interactions Abstract Ticks transmit a variety of viral, bacterial and protozoal pathogens, which are often zoonotic. -

Response of Heterotrophic Stream Biofilm Communities to a Gradient of Resources
The following supplement accompanies the article Response of heterotrophic stream biofilm communities to a gradient of resources D. J. Van Horn1,*, R. L. Sinsabaugh1, C. D. Takacs-Vesbach1, K. R. Mitchell1,2, C. N. Dahm1 1Department of Biology, University of New Mexico, Albuquerque, New Mexico 87131, USA 2Present address: Department of Microbiology & Immunology, University of British Columbia Life Sciences Centre, Vancouver BC V6T 1Z3, Canada *Email: [email protected] Aquatic Microbial Ecology 64:149–161 (2011) Table S1. Representative sequences for each OTU, associated GenBank accession numbers, and taxonomic classifications with bootstrap values (in parentheses), generated in mothur using 14956 reference sequences from the SILVA data base Treatment Accession Sequence name SILVA taxonomy classification number Control JF695047 BF8FCONT18Fa04.b1 Bacteria(100);Proteobacteria(100);Gammaproteobacteria(100);Pseudomonadales(100);Pseudomonadaceae(100);Cellvibrio(100);unclassified; Control JF695049 BF8FCONT18Fa12.b1 Bacteria(100);Proteobacteria(100);Alphaproteobacteria(100);Rhizobiales(100);Methylocystaceae(100);uncultured(100);unclassified; Control JF695054 BF8FCONT18Fc01.b1 Bacteria(100);Planctomycetes(100);Planctomycetacia(100);Planctomycetales(100);Planctomycetaceae(100);Isosphaera(50);unclassified; Control JF695056 BF8FCONT18Fc04.b1 Bacteria(100);Proteobacteria(100);Gammaproteobacteria(100);Xanthomonadales(100);Xanthomonadaceae(100);uncultured(64);unclassified; Control JF695057 BF8FCONT18Fc06.b1 Bacteria(100);Proteobacteria(100);Betaproteobacteria(100);Burkholderiales(100);Comamonadaceae(100);Ideonella(54);unclassified; -

Supplementary Information
doi: 10.1038/nature06269 SUPPLEMENTARY INFORMATION METAGENOMIC AND FUNCTIONAL ANALYSIS OF HINDGUT MICROBIOTA OF A WOOD FEEDING HIGHER TERMITE TABLE OF CONTENTS MATERIALS AND METHODS 2 • Glycoside hydrolase catalytic domains and carbohydrate binding modules used in searches that are not represented by Pfam HMMs 5 SUPPLEMENTARY TABLES • Table S1. Non-parametric diversity estimators 8 • Table S2. Estimates of gross community structure based on sequence composition binning, and conserved single copy gene phylogenies 8 • Table S3. Summary of numbers glycosyl hydrolases (GHs) and carbon-binding modules (CBMs) discovered in the P3 luminal microbiota 9 • Table S4. Summary of glycosyl hydrolases, their binning information, and activity screening results 13 • Table S5. Comparison of abundance of glycosyl hydrolases in different single organism genomes and metagenome datasets 17 • Table S6. Comparison of abundance of glycosyl hydrolases in different single organism genomes (continued) 20 • Table S7. Phylogenetic characterization of the termite gut metagenome sequence dataset, based on compositional phylogenetic analysis 23 • Table S8. Counts of genes classified to COGs corresponding to different hydrogenase families 24 • Table S9. Fe-only hydrogenases (COG4624, large subunit, C-terminal domain) identified in the P3 luminal microbiota. 25 • Table S10. Gene clusters overrepresented in termite P3 luminal microbiota versus soil, ocean and human gut metagenome datasets. 29 • Table S11. Operational taxonomic unit (OTU) representatives of 16S rRNA sequences obtained from the P3 luminal fluid of Nasutitermes spp. 30 SUPPLEMENTARY FIGURES • Fig. S1. Phylogenetic identification of termite host species 38 • Fig. S2. Accumulation curves of 16S rRNA genes obtained from the P3 luminal microbiota 39 • Fig. S3. Phylogenetic diversity of P3 luminal microbiota within the phylum Spirocheates 40 • Fig. -

35 Disorders of Purine and Pyrimidine Metabolism
35 Disorders of Purine and Pyrimidine Metabolism Georges van den Berghe, M.- Françoise Vincent, Sandrine Marie 35.1 Inborn Errors of Purine Metabolism – 435 35.1.1 Phosphoribosyl Pyrophosphate Synthetase Superactivity – 435 35.1.2 Adenylosuccinase Deficiency – 436 35.1.3 AICA-Ribosiduria – 437 35.1.4 Muscle AMP Deaminase Deficiency – 437 35.1.5 Adenosine Deaminase Deficiency – 438 35.1.6 Adenosine Deaminase Superactivity – 439 35.1.7 Purine Nucleoside Phosphorylase Deficiency – 440 35.1.8 Xanthine Oxidase Deficiency – 440 35.1.9 Hypoxanthine-Guanine Phosphoribosyltransferase Deficiency – 441 35.1.10 Adenine Phosphoribosyltransferase Deficiency – 442 35.1.11 Deoxyguanosine Kinase Deficiency – 442 35.2 Inborn Errors of Pyrimidine Metabolism – 445 35.2.1 UMP Synthase Deficiency (Hereditary Orotic Aciduria) – 445 35.2.2 Dihydropyrimidine Dehydrogenase Deficiency – 445 35.2.3 Dihydropyrimidinase Deficiency – 446 35.2.4 Ureidopropionase Deficiency – 446 35.2.5 Pyrimidine 5’-Nucleotidase Deficiency – 446 35.2.6 Cytosolic 5’-Nucleotidase Superactivity – 447 35.2.7 Thymidine Phosphorylase Deficiency – 447 35.2.8 Thymidine Kinase Deficiency – 447 References – 447 434 Chapter 35 · Disorders of Purine and Pyrimidine Metabolism Purine Metabolism Purine nucleotides are essential cellular constituents 4 The catabolic pathway starts from GMP, IMP and which intervene in energy transfer, metabolic regula- AMP, and produces uric acid, a poorly soluble tion, and synthesis of DNA and RNA. Purine metabo- compound, which tends to crystallize once its lism can be divided into three pathways: plasma concentration surpasses 6.5–7 mg/dl (0.38– 4 The biosynthetic pathway, often termed de novo, 0.47 mmol/l). starts with the formation of phosphoribosyl pyro- 4 The salvage pathway utilizes the purine bases, gua- phosphate (PRPP) and leads to the synthesis of nine, hypoxanthine and adenine, which are pro- inosine monophosphate (IMP). -

Microbial Community Response to Heavy and Light Crude Oil in the Great Lakes
Microbial Community Response to Heavy and Light Crude Oil in the Great Lakes Stephen Techtmann 10/24/19 Microbial Sensors Techtmann Lab @ MTU Investigating the applications of environmental microbial communities Hydraulic Fracturing Related Antibiotic Resistance Oil Bioremediation Techtmann Lab @ MTU Overview • Background on oil biodegradation • Microbial response to light and heavy crude oil in the Great Lakes • Machine learning for prediction of contamination in the Great Lakes. Oil Spills Deepwater Horizon Enbridge Line 6B Deepwater Horizon Oil Spill • 4,1000,000 bbl of oil released • Light Sweet Crude oil released • April 20, 2010 • 1101.7 miles of shoreline oiled Atlas and Hazen 2011 Enbridge Line 6B Spill – Marshall MI • 20,082 bbl of oil released • Diluted Bitumen • July 26, 2010 • 70 miles of shoreline oiled https://www.mlive.com/news/kalamazoo/2010/07/state_of_emergency_declared_as.html Oil Transmissions Pipelines in the Great Lakes Region Line 5: • 645 miles from Superior WI to Sarnia Ontario • 540,000 barrels per day • Light crude and natural gas liquids (NGLs) Crude oil Oil types and API Gravity Microbes and Biotechnology (Bioremediation) Low cost input Microbe High value output Decreased Cost Contaminant Increased Efficiency Carbon dioxide or non- toxic daughter products Carbon dioxide Microbial Biomass Petroleum Microbe Daughter Products Water Microbial Ecology and Biotechnology Low cost input Microbe High value output Decreased Cost/Increased Efficiency Complex input Input A Microbe Microbe Output A Input B Microbe Output -

Downloaded 13 April 2017); Using Diamond
bioRxiv preprint doi: https://doi.org/10.1101/347021; this version posted June 14, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 1 2 3 4 5 Re-evaluating the salty divide: phylogenetic specificity of 6 transitions between marine and freshwater systems 7 8 9 10 Sara F. Pavera, Daniel J. Muratorea, Ryan J. Newtonb, Maureen L. Colemana# 11 a 12 Department of the Geophysical Sciences, University of Chicago, Chicago, Illinois, USA 13 b School of Freshwater Sciences, University of Wisconsin Milwaukee, Milwaukee, Wisconsin, USA 14 15 Running title: Marine-freshwater phylogenetic specificity 16 17 #Address correspondence to Maureen Coleman, [email protected] 18 bioRxiv preprint doi: https://doi.org/10.1101/347021; this version posted June 14, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 19 Abstract 20 Marine and freshwater microbial communities are phylogenetically distinct and transitions 21 between habitat types are thought to be infrequent. We compared the phylogenetic diversity of 22 marine and freshwater microorganisms and identified specific lineages exhibiting notably high or 23 low similarity between marine and freshwater ecosystems using a meta-analysis of 16S rRNA 24 gene tag-sequencing datasets. As expected, marine and freshwater microbial communities 25 differed in the relative abundance of major phyla and contained habitat-specific lineages; at the 26 same time, however, many shared taxa were observed in both environments. 27 Betaproteobacteria and Alphaproteobacteria sequences had the highest similarity between 28 marine and freshwater sample pairs. -

1 Supplementary Information Ugly Ducklings – the Dark Side of Plastic
Supplementary Information Ugly ducklings – The dark side of plastic materials in contact with potable water Lisa Neu1,2, Carola Bänziger1, Caitlin R. Proctor1,2, Ya Zhang3, Wen-Tso Liu3, Frederik Hammes1,* 1 Eawag, Swiss Federal Institute of Aquatic Science and Technology, Dübendorf, Switzerland 2 Department of Environmental Systems Science, Institute of Biogeochemistry and Pollutant Dynamics, ETH Zürich, Zürich, Switzerland 3 Department of Civil and Environmental Engineering, University of Illinois at Urbana-Champaign, USA Table of contents Table S1 Exemplary online blog entries on biofouling in bath toys Figure S1 Images of all examined bath toys Figure S2 Additional images of bath toy biofilms by OCT Figure S3 Additional images on biofilm composition by SEM Figure S4 Number of bacteria and proportion of intact cells in bath toy biofilms Table S2 Classification of shared OTUs between bath toys Table S3 Shared and ‘core’ communities in bath toys from single households Table S4 Richness and diversity Figure S5 Classification of abundant OTUs in real bath toy biofilms Table S5 Comparison of most abundant OTUs in control bath toy biofilms Figure S6 Fungal community composition in bath toy biofilms Table S6 Conventional plating results for indicator bacteria and groups Table S7 Bioavailability of migrating carbon from control bath toys’ material Water chemistry Method and results (Table S8) Table S9 Settings for Amplification PCR and Index PCR reactions 1 Table S1: Exemplary online blog entries on biofouling inside bath toys Issue - What is the slime? Link Rub-a-dub-dub, https://www.babble.com/baby/whats-in-the-tub/ what’s in the tub? What’s the black stuff http://blogs.babycenter.com/momstories/whats-the-black- in your squeeze toys? stuff-in-your-squeeze-toys/ Friday Find: NBC’s http://www.bebravekeepgoing.com/2010/03/friday-find-nbcs- Today Show segment: today-show-segment-do.html Do bath toys carry germs? Yuck. -

Examining the Antimicrobial Activity of Cefepime-Taniborbactam
Examining the antimicrobial activity of cefepime-taniborbactam (formerly cefepime/VNRX-5133) against Burkholderia species isolated from cystic fibrosis patients in the United States Elise T. Zeiser1, Scott A. Becka1, John J. LiPuma2, David A. Six 3, Greg Moeck 3, and Krisztina M. Papp-Wallace1,4 1Veterans Affairs Northeast Ohio Healthcare System, Cleveland, OH; 2University of Michigan, Ann Arbor, MI; 3Venatorx 4 Pharmaceuticals, Inc., Malvern, PA; and Case Western Reserve University, Cleveland, OH Correspondence to: [email protected] Abstract Results Background: Burkholderia cepacia complex (Bcc), a group of >20 related species, and B. gladioli are 60 opportunistic human pathogens that cause chronic infections in people with cystic fibrosis (CF) or cefepime compromised immune systems. Ceftazidime and trimethoprim-sulfamethoxazole are first-line agents used to treat infections due to Burkholderia spp. However, these species have developed resistance to cefepime-taniborbactam many antibiotics, including first-line therapies. β-lactam resistance in Burkholderia species is largely 40 mediated by PenA-like chromosomal class A β-lactamases. A novel investigational β-lactam/β-lactamase inhibitor combination, cefepime-taniborbactam (formerly cefepime/VNRX-5133) demonstrates potent antimicrobial activity against gram-negative bacteria producing class A, B, C, and D β-lactamases. The activity of cefepime-taniborbactam was investigated against Bcc and B. gladioli; moreover, the biochemical activity of taniborbactam against the PenA1 carbapenemase was evaluated. 20 Methods: CLSI-based agar dilution antimicrobial susceptibility testing using cefepime and cefepime combined with taniborbactam at 4 mg/L was conducted against a curated panel of 150 Burkholderia species obtained from the Burkholderia cepacia Research Laboratory and Repository. Isolates were Number isolates of recovered from respiratory specimens from 150 different individuals with CF receiving care in 68 cities 0 throughout 36 states within the United States.