1 the Trigeminal Nerve. Part I: an Over-View
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Perioral Gustatory Sweating: Case Report
The Journal of Laryngology & Otology (2012), 126, 532–534. CLINICAL RECORD © JLO (1984) Limited, 2012 doi:10.1017/S0022215112000229 Perioral gustatory sweating: case report S C KÄYSER1, K J A O INGELS2, F J A VAN DEN HOOGEN2 1Department of Primary and Community Care, Radboud University Nijmegen Medical Centre, and 2Department of Otorhinolaryngology/Head and Neck Surgery, Radboud University Nijmegen Medical Centre, The Netherlands Abstract Objective: Presentation of a case of perioral Frey syndrome. Design: Case report. Subject: A 72-year-old woman with hyperhidrosis around the mouth and chin. Results: This patient suffered from bilateral perioral gustatory sweating following a mandibular osteotomy; such a case has not previously been described. Possible pathophysiological hypotheses are discussed in relation to the anatomy and innervation of the salivary glands. Conclusion: Perioral gustatory sweating is a rare complication of osteotomy. Key words: Gustatory sweating; Frey Syndrome; Perioral; Hyperhidrosis Introduction perioral excessive sweating and flushing which only Frey syndrome, also known as auriculotemporal syndrome, is a occurred during (and not preceding) eating. Her complaints well-known complication of parotid surgery. Approximately had begun following bilateral osteotomy of the mandible in 24 per cent of patients undergoing parotidectomy experience 1960, at the age of 25 years. The indication for this procedure gustatory sweating, although the reported incidence varies had been prognathism. Her recovery had been complicated greatly.1 Frey syndrome appears following a latency period by inadequate bone healing and loss of the right and left of one to 36 months (or longer) after surgery.2,3 inferior alveolar nerves (Figure 1). The aetiology of Frey syndrome is explained by ‘aberrant The diagnosis of hyperhidrosis was made using the nervous regeneration’. -

The Morphometry of the Angle of Mandible and Its Correlation with Age and Sex in the Ethekwini Metropolitan Region: a Panoramic Study
Int. J. Morphol., 35(2):661-666, 2017. The Morphometry of the Angle of Mandible and its Correlation with Age and Sex in the eThekwini Metropolitan Region: A Panoramic Study Morfometría del Angulo de la Mandíbula y su Correlación con Edad y Sexo en la Región Metropolitana eThekwini: Un Estudio Panorámico S. Pillay1; S. Ishwarkumar1; B.Z. De Gama1 & P. Pillay1 PILLAY, S.; ISHWARKUMAR, S.; DE GAMA, B. Z. & PILLAY, P. The morphometry of the angle of mandible and its correlation with age and sex in the eThekwini metropolitan region: A panoramic study. Int. J. Morphol., 35(2):661-666, 2017. SUMMARY: The angle of mandible is formed by the tangent line joining the posterior margin of the ramus and the base of the mandible. The angle of mandible has population-specific characteristics therefore; it is imperative to the field of forensic anthropology for age and sex determination. Literary reports regarding the use of the angle of mandible for age and sex determination vary, as some studies support it, while other studies have documented inefficiencies. Therefore, the aim of this investigation was to document the morphometry of the angle of mandible and to determine if a correlation between the angle of mandible, age and sex exists. Sixty four digital panoramic radiographs (n=128) of individuals between 16-30 years were morphometrically analysed using the Dicom Digital Imaging Software. The data was captured and analysed using the Statistical Package for Social Science (SPSS version 23.0). Despite females having a greater angle of mandible than males, no statistically significant correlation was found between the size of the angle of mandible and sex (p=0.088). -
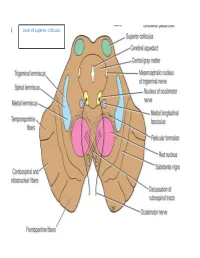
Level of Superior Colliculus ° Edinger- Westphal Nucleus ° Pretectal Nucleus: Close to the Lateral Part of the Superior Colliculus
Level of superior colliculus ° Edinger- Westphal nucleus ° pretectal nucleus: close to the lateral part of the superior colliculus. Red nucleus • Rounded mass of gray matter • Situated bt cerebral aqueduct and substantia nigra • Reddish blue(vascularity & iron containing pigment) • Afferents from: cerebral cortex,cerebellum,substa ntia nigra, thalamic nuclei, spinal cord • Efferent to: spinal cord, reticular formation. thalamus and substantia nigra • involved in motor coordination. Crus cerebri • Corticospinal & corticonuclear fibers (middle) • Frontopontine fibers (medial) • Temporopontine fibers (lateral) these descending tracts connect the cerebral cortex with spinal cord, cranial nerves nuclei, pons & cerebellum Level at superior colliculus ° Superior colliculus ° Occulomotor nucleus (posterior to MLF) ° Occulomotor n emerges through red nucleus ° Edinger-Westphal nucleus ° pretectal nucleus: close to the lateral part of the superior colliculus. ° MLF ° Medial , trigeminal, spinal leminiscus ( no lateral leminiscus) ° Red nucleus ° Substantia nigra ° Crus cerebri ° RF Substantia nigra ° Large motor nucleus ° is a brain structure located in the midbrain ° plays an important role in reward, addiction, and movement. ° Substantia nigra is Latin for "black substance" due to high levels of melanin ° has connections with basal ganglia ,cerebral cortex ° Concerned with muscle tone ° Parkinson's disease is caused by the death of neurons in the substantia nigra Oculomotor Nerve (III) • Main oculomotor nucleus • Accessory parasympathetic nucleus -

The Gonial Angle Relation with Different Dentitiontal Status on Orthopantomogram in Iraqi Sample
Republic of Iraq Ministry of Higher Education And scientific Research University of Baghdad College of Dentistry The Gonial angle relation with different dentitiontal status on orthopantomogram in iraqi sample A project Submitted to Collage of Dentistry, University of Baghdad. Department of Oral and Maxillofacial Radiology in fulfillment for the requirement to award the degree B.D.S Done by Hadeer Majid Ali 5th Grade Supervisor Dr. Alaa Salah Mahdi B.D.S, M.SC. Oral and Maxillofacial Radiology Baghdad Iraq 2018_1439 Dedication To my parents who were their for me in every step of the way with their have love and support… To my supervisor for his guidance, help and endless support throughout this project… Hadeer Majid Ali Abstract Abstract Background: Mandibular angle plays an important role in ensuring a harmonious facial profile from esthetic point of view so it is a representative of mandible morphology. Resorption of alveolar bone is the best recognized feature of mandibular aging in the edentate subjects and changes of the mandibular cortical shape and thickness may be used as indications to many abnormalities, such as osteoporosis. Panoramic radiographs are a useful tool for the measurement because majority of dentists request an Orthopantomogram for patients during routine dental examination. The Aim of the study: to correlated the gonial angle relation with different dentitional status they are in three group dentulous, partial dentulous and edentulous using digital panoramic imaging system with age, gender and dental status. Subjects, Materials and Methods: This study was conducted on 30 Iraqi in three group dentulous , partial dentulous and edentulous attending to the digital panoramic clinic of the hospital of college of dentistry university of Baghdad Information from each subject was recorded in a special case sheet. -

Cranial Nerve Palsy
Cranial Nerve Palsy What is a cranial nerve? Cranial nerves are nerves that lead directly from the brain to parts of our head, face, and trunk. There are 12 pairs of cranial nerves and some are involved in special senses (sight, smell, hearing, taste, feeling) while others control muscles and glands. Which cranial nerves pertain to the eyes? The second cranial nerve is called the optic nerve. It sends visual information from the eye to the brain. The third cranial nerve is called the oculomotor nerve. It is involved with eye movement, eyelid movement, and the function of the pupil and lens inside the eye. The fourth cranial nerve is called the trochlear nerve and the sixth cranial nerve is called the abducens nerve. They each innervate an eye muscle involved in eye movement. The fifth cranial nerve is called the trigeminal nerve. It provides facial touch sensation (including sensation on the eye). What is a cranial nerve palsy? A palsy is a lack of function of a nerve. A cranial nerve palsy may cause a complete or partial weakness or paralysis of the areas served by the affected nerve. In the case of a cranial nerve that has multiple functions (such as the oculomotor nerve), it is possible for a palsy to affect all of the various functions or only some of the functions of that nerve. What are some causes of a cranial nerve palsy? A cranial nerve palsy can occur due to a variety of causes. It can be congenital (present at birth), traumatic, or due to blood vessel disease (hypertension, diabetes, strokes, aneurysms, etc). -

Anatomy of Maxillary and Mandibular Local Anesthesia
Anatomy of Mandibular and Maxillary Local Anesthesia Patricia L. Blanton, Ph.D., D.D.S. Professor Emeritus, Department of Anatomy, Baylor College of Dentistry – TAMUS and Private Practice in Periodontics Dallas, Texas Anatomy of Mandibular and Maxillary Local Anesthesia I. Introduction A. The anatomical basis of local anesthesia 1. Infiltration anesthesia 2. Block or trunk anesthesia II. Review of the Trigeminal Nerve (Cranial n. V) – the major sensory nerve of the head A. Ophthalmic Division 1. Course a. Superior orbital fissure – root of orbit – supraorbital foramen 2. Branches – sensory B. Maxillary Division 1. Course a. Foramen rotundum – pterygopalatine fossa – inferior orbital fissure – floor of orbit – infraorbital 2. Branches - sensory a. Zygomatic nerve b. Pterygopalatine nerves [nasal (nasopalatine), orbital, palatal (greater and lesser palatine), pharyngeal] c. Posterior superior alveolar nerves d. Infraorbital nerve (middle superior alveolar nerve, anterior superior nerve) C. Mandibular Division 1. Course a. Foramen ovale – infratemporal fossa – mandibular foramen, Canal -> mental foramen 2. Branches a. Sensory (1) Long buccal nerve (2) Lingual nerve (3) Inferior alveolar nerve -> mental nerve (4) Auriculotemporal nerve b. Motor (1) Pterygoid nerves (2) Temporal nerves (3) Masseteric nerves (4) Nerve to tensor tympani (5) Nerve to tensor veli palatine (6) Nerve to mylohyoid (7) Nerve to anterior belly of digastric c. Both motor and sensory (1) Mylohyoid nerve III. Usual Routes of innervation A. Maxilla 1. Teeth a. Molars – Posterior superior alveolar nerve b. Premolars – Middle superior alveolar nerve c. Incisors and cuspids – Anterior superior alveolar nerve 2. Gingiva a. Facial/buccal – Superior alveolar nerves b. Palatal – Anterior – Nasopalatine nerve; Posterior – Greater palatine nerves B. -

The Sympathetic and the Parasympathetic Nervous System
The sympathetic and the parasympathetic nervous system Zsuzsanna Tóth, PhD Institute of Anatomy, Histology and Embryology Semmelweis University The role of the autonomic nervous system Claude Bernard • „milieu intérieur” concept; every organism lives in its internal environment that is constant and independent form the external environment Walter Bradford Cannon homeostasis; • an extension of the “milieu interieur” concept • consistence in an open system requires mechanisms that act to maintain that consistency • steady-state conditions require that any tendency toward change automatically meets with factors that resist that change • regulating systems that determine the homeostatic state : o autonomic nervous system ( sympathetic, parasympathetic, enteral) o endocrine system General structure of the autonomic nervous system craniosacral thoracolumbar Anatomy Neurotransmittersof the gut autonomic nervous system. symp. gangl pregangl. fiber pregangl. postgangl. fiber fiber (PoR) PoR enteral ganglion PoR PoR smooth muscle smooth muscle Kuratani S Development 2009;136:1585-1589 Sympathetic activation: Fight or flight reaction • energy mobilization • preparation for escape, or fight vasoconstriction • generalized Parasympathetic activation: adrenal • energy saving and restoring • „rest and digest” system • more localized vasoconstriction Paravertebral ganglia and the sympathetic chains pars cervicalis superius ganglion medium cervicale stellatum pars vertebrae • from the base of the skull to the caudal end thoracalis thoracalis of the sacrum • paravertebral ganglia (ganglia trunci sympathici) • rami interganglionares pars vertebrae • the two chains fuses at the ganglion impar abdominalis lumbalis sacrum pars pelvina foramen sacralia anteriora ganglion impar Anatomy of the cervical part of the sympathetic trunk superior cervical ganglion • behind the seath of the carotid, fusiform ggl. cervicale superius • IML T1-3 vegetative motoneurons- preganglionic fibers truncus symp. -

Simple Ways to Dissect Ciliary Ganglion for Orbital Anatomical Education
OkajimasDetection Folia Anat. of ciliary Jpn., ganglion94(3): 119–124, for orbit November, anatomy 2017119 Simple ways to dissect ciliary ganglion for orbital anatomical education By Ming ZHOU, Ryoji SUZUKI, Hideo AKASHI, Akimitsu ISHIZAWA, Yoshinori KANATSU, Kodai FUNAKOSHI, Hiroshi ABE Department of Anatomy, Akita University Graduate School of Medicine, Akita, 010-8543 Japan –Received for Publication, September 21, 2017– Key Words: ciliary ganglion, orbit, human anatomy, anatomical education Summary: In the case of anatomical dissection as part of medical education, it is difficult for medical students to find the ciliary ganglion (CG) since it is small and located deeply in the orbit between the optic nerve and the lateral rectus muscle and embedded in the orbital fat. Here, we would like to introduce simple ways to find the CG by 1): tracing the sensory and parasympathetic roots to find the CG from the superior direction above the orbit, 2): transecting and retracting the lateral rectus muscle to visualize the CG from the lateral direction of the orbit, and 3): taking out whole orbital structures first and dissecting to observe the CG. The advantages and disadvantages of these methods are discussed from the standpoint of decreased laboratory time and students as beginners at orbital anatomy. Introduction dissection course for the first time and with limited time. In addition, there are few clear pictures in anatomical The ciliary ganglion (CG) is one of the four para- textbooks showing the morphology of the CG. There are sympathetic ganglia in the head and neck region located some scientific articles concerning how to visualize the behind the eyeball between the optic nerve and the lateral CG, but they are mostly based on the clinical approaches rectus muscle in the apex of the orbit (Siessere et al., rather than based on the anatomical procedure for medical 2008). -

Morfofunctional Structure of the Skull
N.L. Svintsytska V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 Ministry of Public Health of Ukraine Public Institution «Central Methodological Office for Higher Medical Education of MPH of Ukraine» Higher State Educational Establishment of Ukraine «Ukranian Medical Stomatological Academy» N.L. Svintsytska, V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 2 LBC 28.706 UDC 611.714/716 S 24 «Recommended by the Ministry of Health of Ukraine as textbook for English- speaking students of higher educational institutions of the MPH of Ukraine» (minutes of the meeting of the Commission for the organization of training and methodical literature for the persons enrolled in higher medical (pharmaceutical) educational establishments of postgraduate education MPH of Ukraine, from 02.06.2016 №2). Letter of the MPH of Ukraine of 11.07.2016 № 08.01-30/17321 Composed by: N.L. Svintsytska, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor V.H. Hryn, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor This textbook is intended for undergraduate, postgraduate students and continuing education of health care professionals in a variety of clinical disciplines (medicine, pediatrics, dentistry) as it includes the basic concepts of human anatomy of the skull in adults and newborns. Rewiewed by: O.M. Slobodian, Head of the Department of Anatomy, Topographic Anatomy and Operative Surgery of Higher State Educational Establishment of Ukraine «Bukovinian State Medical University», Doctor of Medical Sciences, Professor M.V. -
The Mandibular Nerve - Vc Or VIII by Prof
The Mandibular Nerve - Vc or VIII by Prof. Dr. Imran Qureshi The Mandibular nerve is the third and largest division of the trigeminal nerve. It is a mixed nerve. Its sensory root emerges from the posterior region of the semilunar ganglion and is joined by the motor root of the trigeminal nerve. These two nerve bundles leave the cranial cavity through the foramen ovale and unite immediately to form the trunk of the mixed mandibular nerve that passes into the infratemporal fossa. Here, it runs anterior to the middle meningeal artery and is sandwiched between the superior head of the lateral pterygoid and tensor veli palatini muscles. After a short course during which a meningeal branch to the dura mater, and the nerve to part of the medial pterygoid muscle (and the tensor tympani and tensor veli palatini muscles) are given off, the mandibular trunk divides into a smaller anterior and a larger posterior division. The anterior division receives most of the fibres from the motor root and distributes them to the other muscles of mastication i.e. the lateral pterygoid, medial pterygoid, temporalis and masseter muscles. The nerve to masseter and two deep temporal nerves (anterior and posterior) pass laterally above the medial pterygoid. The nerve to the masseter continues outward through the mandibular notch, while the deep temporal nerves turn upward deep to temporalis for its supply. The sensory fibres that it receives are distributed as the buccal nerve. The 1 | P a g e buccal nerve passes between the medial and lateral pterygoids and passes downward and forward to emerge from under cover of the masseter with the buccal artery. -

An Unusual Finding of the Auriculotemporal Nerve: Possible Risk Factor During Preauricular Skin Incisions
Case Report An unusual finding of the auriculotemporal nerve: possible risk factor during preauricular skin incisions Joe Iwanaga1,2,3, Samuel L. Bobek4, Christian Fisahn1,5, Ken Nakamura3, Yoshihiro Miyazono3, R. Shane Tubbs1 1Seattle Science Foundation, Seattle, WA 98122, USA; 2Department of Anatomy, 3Dental and Oral Medical Center, Kurume University School of Medicine, Kurume, Fukuoka 830-0011, Japan; 4Swedish Maxillofacial Surgery, 5Swedish Neuroscience Institute, Swedish Medical Center, Seattle, WA 98122, USA Correspondence to: Joe Iwanaga. Seattle Science Foundation, 550 17th Ave, James Tower, Suite 600, Seattle, WA 98122, USA. Email: [email protected]. Abstract: The auriculotemporal nerve (ATN) is a branch of the mandibular nerve and has been implicated for some migraines and its role in Frey’s syndrome is well known. An adult cadaver was found to have a duplicated ATN. The anterior trunk ascended as the superficial temporal artery and gave off the branches to the temporomandibular joint, parotid gland, external acoustic meatus and temporal region and communicated with a posterior trunk of the ATN. The posterior trunk ascended via the subcutaneous tissues 1 mm anterior to the auricle and gave off the branches to the anterior auricular region, temporal region and communicated with the anterior trunk. Such a duplicated ATN might be injured with preauricular skin incisions. Knowledge of such an anatomical variation might assist surgeons in iatrogenic injury of the ATN. Keywords: Auriculotemporal nerve (ATN); infratemporal fossa; Frey’s syndrome; mandibular nerve Submitted Aug 09, 2016. Accepted for publication Aug 17, 2016. doi: 10.21037/gs.2016.09.02 View this article at: http://dx.doi.org/10.21037/gs.2016.09.02 Introduction Case presentation The auriculotemporal nerve (ATN) is one of the branches During the dissection of a cadaver that was 87-year-old at of the mandibular division (V3) of the trigeminal nerve. -

PERFORATION of INFERIOR ALVEOLAR NERVE by MAXILLARY ARTERY Prakash Billakanti Babu, Ramachandra Bhat K, Vanishree S Nayak
Inferior alveolar nerve Rev Arg de Anat Clin; 2011, 3 (3): 124-129 __________________________________________________________________________________________ Case report PERFORATION OF INFERIOR ALVEOLAR NERVE BY MAXILLARY ARTERY Prakash Billakanti Babu, Ramachandra Bhat K, Vanishree S Nayak Kasturba Medical College, Manipal University, Manipal, Karnataka, India RESUMEN La fosa infratemporal es un área anatómica inferior alveolar nerve originated from the clínicamente importante para la administración de mandibular nerve by two roots and the first part agentes anestésicos locales en odontología y cirugía of the maxillary artery was incorporated between maxilofacial. Fueron estudiadas variaciones en la them. An embryologic origin of this variation and anatomía del nervio alveolar inferior y la arteria maxilar its clinical implications is discussed. Because the en la disección infratemporal. Durante la disección rutinaria de la cabeza en el cadáver de un varón maxillary artery runs between the two roots of the adulto, fue observada una variación excepcional en el inferior alveolar nerve, and the nerve was fixed origen del nervio alveolar inferior y su relación con las between the foramen ovale and mandibular estructuras circundantes. El nervio alveolar inferior se foramen, neurovascular entrapment may cause originaba en el nervio mandibular por dos raíces y la pain numbness or headache and may interfere primera parte de la arteria maxilar estaba incorporada with the injection of local anesthetics into the entre ambas. El origen embriológico de esta variación infratemporal fossa. Anatomical variations in this y sus implicaciones clínicas es debatido. Dado que la region should be kept in mind, particularly in arteria maxilar transcurría entre las dos raíces del cases of failed treatment of trigeminal neuralgia.