Invasive Cerambycid Pests and Biosecurity Measures. Chapter
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Green-Tree Retention and Controlled Burning in Restoration and Conservation of Beetle Diversity in Boreal Forests
Dissertationes Forestales 21 Green-tree retention and controlled burning in restoration and conservation of beetle diversity in boreal forests Esko Hyvärinen Faculty of Forestry University of Joensuu Academic dissertation To be presented, with the permission of the Faculty of Forestry of the University of Joensuu, for public criticism in auditorium C2 of the University of Joensuu, Yliopistonkatu 4, Joensuu, on 9th June 2006, at 12 o’clock noon. 2 Title: Green-tree retention and controlled burning in restoration and conservation of beetle diversity in boreal forests Author: Esko Hyvärinen Dissertationes Forestales 21 Supervisors: Prof. Jari Kouki, Faculty of Forestry, University of Joensuu, Finland Docent Petri Martikainen, Faculty of Forestry, University of Joensuu, Finland Pre-examiners: Docent Jyrki Muona, Finnish Museum of Natural History, Zoological Museum, University of Helsinki, Helsinki, Finland Docent Tomas Roslin, Department of Biological and Environmental Sciences, Division of Population Biology, University of Helsinki, Helsinki, Finland Opponent: Prof. Bengt Gunnar Jonsson, Department of Natural Sciences, Mid Sweden University, Sundsvall, Sweden ISSN 1795-7389 ISBN-13: 978-951-651-130-9 (PDF) ISBN-10: 951-651-130-9 (PDF) Paper copy printed: Joensuun yliopistopaino, 2006 Publishers: The Finnish Society of Forest Science Finnish Forest Research Institute Faculty of Agriculture and Forestry of the University of Helsinki Faculty of Forestry of the University of Joensuu Editorial Office: The Finnish Society of Forest Science Unioninkatu 40A, 00170 Helsinki, Finland http://www.metla.fi/dissertationes 3 Hyvärinen, Esko 2006. Green-tree retention and controlled burning in restoration and conservation of beetle diversity in boreal forests. University of Joensuu, Faculty of Forestry. ABSTRACT The main aim of this thesis was to demonstrate the effects of green-tree retention and controlled burning on beetles (Coleoptera) in order to provide information applicable to the restoration and conservation of beetle species diversity in boreal forests. -

Pharmaceutical Sciences
IAJPS 2019, 06 (10), 12713-12727 Muhammad Yousaf Ghilzai et al ISSN 2349-7750 CODEN [USA]: IAJPBB ISSN: 2349-7750 INDO AMERICAN JOURNAL OF PHARMACEUTICAL SCIENCES Available online at: http://www.iajps.com Research Article FOOD PREFERENCE OF QUETTA BORER, AEOLESTHES SARTA COLEOPTERA; (CERAMBYCIDAE) UNDER CONTROL CONDITION 1Muhammad Yousaf Ghilzai,1Arshad Ghani Khan,2Imran Ali Sani, 2Nisar Ahmed 1Mohammad Amin 1Zafarullah,1Azhar Sheikh,1Essa Khan, 1Zia ul Haq,2Umair Ahmed, 3Sajid Nabi 1Balochistan Agriculture College Quetta, Pakistan 2Balochistan University of Information Technology Engineering and Management sciences, BUITEMS, Quetta, Pakistan 3University of Balochistan, Quetta Pakistan Article Received: March 2019 Accepted: April 2019 Published: October 2019 Abstract: Balochistan is the largest province of Pakistan, which is blessed with four agro-ecological zones and it has unique environment condition for the production of a great variety of quality fruits, that’s why the province is known as the fruit-basket of the country. Among these fruits’ apple, apricot and almond are considered as important fruits of the world and Pakistan. Fruits provide nutrients and vitamins to the human. These fruits are generally grown in temperate regions of Pakistan especially in Balochistan and Khyber Pakhtunkhwa (KPK). These fruits are attacked by number of insect pests viz, codling moth, shot hole borer, aphids and san Jose scale but borers (roundhead borer and flat-headed borer) are most serious/ destructive pest and potential threat to deciduous fruit of Balochistan. Adult stages feed on the buds, cortex and causing heavy leaf defoliation. While, immature stage (larvae) bore into the tree and destroyed their xylem and phloem bundle which badly affects the yield of the tree. -

4 Reproductive Biology of Cerambycids
4 Reproductive Biology of Cerambycids Lawrence M. Hanks University of Illinois at Urbana-Champaign Urbana, Illinois Qiao Wang Massey University Palmerston North, New Zealand CONTENTS 4.1 Introduction .................................................................................................................................. 133 4.2 Phenology of Adults ..................................................................................................................... 134 4.3 Diet of Adults ............................................................................................................................... 138 4.4 Location of Host Plants and Mates .............................................................................................. 138 4.5 Recognition of Mates ................................................................................................................... 140 4.6 Copulation .................................................................................................................................... 141 4.7 Larval Host Plants, Oviposition Behavior, and Larval Development .......................................... 142 4.8 Mating Strategy ............................................................................................................................ 144 4.9 Conclusion .................................................................................................................................... 148 Acknowledgments ................................................................................................................................. -

Hymenoptera: Eulophidae) 321-356 ©Entomofauna Ansfelden/Austria; Download Unter
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Entomofauna Jahr/Year: 2007 Band/Volume: 0028 Autor(en)/Author(s): Yefremova Zoya A., Ebrahimi Ebrahim, Yegorenkova Ekaterina Artikel/Article: The Subfamilies Eulophinae, Entedoninae and Tetrastichinae in Iran, with description of new species (Hymenoptera: Eulophidae) 321-356 ©Entomofauna Ansfelden/Austria; download unter www.biologiezentrum.at Entomofauna ZEITSCHRIFT FÜR ENTOMOLOGIE Band 28, Heft 25: 321-356 ISSN 0250-4413 Ansfelden, 30. November 2007 The Subfamilies Eulophinae, Entedoninae and Tetrastichinae in Iran, with description of new species (Hymenoptera: Eulophidae) Zoya YEFREMOVA, Ebrahim EBRAHIMI & Ekaterina YEGORENKOVA Abstract This paper reflects the current degree of research of Eulophidae and their hosts in Iran. A list of the species from Iran belonging to the subfamilies Eulophinae, Entedoninae and Tetrastichinae is presented. In the present work 47 species from 22 genera are recorded from Iran. Two species (Cirrospilus scapus sp. nov. and Aprostocetus persicus sp. nov.) are described as new. A list of 45 host-parasitoid associations in Iran and keys to Iranian species of three genera (Cirrospilus, Diglyphus and Aprostocetus) are included. Zusammenfassung Dieser Artikel zeigt den derzeitigen Untersuchungsstand an eulophiden Wespen und ihrer Wirte im Iran. Eine Liste der für den Iran festgestellten Arten der Unterfamilien Eu- lophinae, Entedoninae und Tetrastichinae wird präsentiert. Mit vorliegender Arbeit werden 47 Arten in 22 Gattungen aus dem Iran nachgewiesen. Zwei neue Arten (Cirrospilus sca- pus sp. nov. und Aprostocetus persicus sp. nov.) werden beschrieben. Eine Liste von 45 Wirts- und Parasitoid-Beziehungen im Iran und ein Schlüssel für 3 Gattungen (Cirro- spilus, Diglyphus und Aprostocetus) sind in der Arbeit enthalten. -

(Coleoptera) of Peru Miguel A
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Center for Systematic Entomology, Gainesville, Insecta Mundi Florida 2-29-2012 Preliminary checklist of the Cerambycidae, Disteniidae, and Vesperidae (Coleoptera) of Peru Miguel A. Monné Universidade Federal do Rio de Janeiro, [email protected] Eugenio H. Nearns University of New Mexico, [email protected] Sarah C. Carbonel Carril Universidad Nacional Mayor de San Marcos, Peru, [email protected] Ian P. Swift California State Collection of Arthropods, [email protected] Marcela L. Monné Universidade Federal do Rio de Janeiro, [email protected] Follow this and additional works at: http://digitalcommons.unl.edu/insectamundi Part of the Entomology Commons Monné, Miguel A.; Nearns, Eugenio H.; Carbonel Carril, Sarah C.; Swift, Ian P.; and Monné, Marcela L., "Preliminary checklist of the Cerambycidae, Disteniidae, and Vesperidae (Coleoptera) of Peru" (2012). Insecta Mundi. Paper 717. http://digitalcommons.unl.edu/insectamundi/717 This Article is brought to you for free and open access by the Center for Systematic Entomology, Gainesville, Florida at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Insecta Mundi by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. INSECTA MUNDI A Journal of World Insect Systematics 0213 Preliminary checklist of the Cerambycidae, Disteniidae, and Vesperidae (Coleoptera) of Peru Miguel A. Monné Museu Nacional Universidade Federal do Rio de Janeiro Quinta da Boa Vista São Cristóvão, 20940-040 Rio de Janeiro, RJ, Brazil Eugenio H. Nearns Department of Biology Museum of Southwestern Biology University of New Mexico Albuquerque, NM 87131-0001, USA Sarah C. Carbonel Carril Departamento de Entomología Museo de Historia Natural Universidad Nacional Mayor de San Marcos Avenida Arenales 1256, Lima, Peru Ian P. -

Longhorn Beetles (Coleoptera, Cerambycidae) Christian Cocquempot, Ake Lindelöw
Longhorn beetles (Coleoptera, Cerambycidae) Christian Cocquempot, Ake Lindelöw To cite this version: Christian Cocquempot, Ake Lindelöw. Longhorn beetles (Coleoptera, Cerambycidae). Alien terrestrial arthropods of Europe, 4 (1), Pensoft Publishers, 2010, BioRisk, 978-954-642-554-6. 10.3897/biorisk.4.56. hal-02823535 HAL Id: hal-02823535 https://hal.inrae.fr/hal-02823535 Submitted on 6 Jun 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. A peer-reviewed open-access journal BioRisk 4(1): 193–218 (2010)Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1 193 doi: 10.3897/biorisk.4.56 RESEARCH ARTICLE BioRisk www.pensoftonline.net/biorisk Longhorn beetles (Coleoptera, Cerambycidae) Chapter 8.1 Christian Cocquempot1, Åke Lindelöw2 1 INRA UMR Centre de Biologie et de Gestion des Populations, CBGP, (INRA/IRD/CIRAD/Montpellier SupAgro), Campus international de Baillarguet, CS 30016, 34988 Montférrier-sur-Lez, France 2 Swedish university of agricultural sciences, Department of ecology. P.O. Box 7044, S-750 07 Uppsala, Sweden Corresponding authors: Christian Cocquempot ([email protected]), Åke Lindelöw (Ake.Linde- [email protected]) Academic editor: David Roy | Received 28 December 2009 | Accepted 21 May 2010 | Published 6 July 2010 Citation: Cocquempot C, Lindelöw Å (2010) Longhorn beetles (Coleoptera, Cerambycidae). -

Bark Beetle Pheromones and Pine Volatiles: Attractant Kairomone Lure Blend for Longhorn Beetles (Cerambycidae) in Pine Stands of the Southeastern United States
FOREST ENTOMOLOGY Bark Beetle Pheromones and Pine Volatiles: Attractant Kairomone Lure Blend for Longhorn Beetles (Cerambycidae) in Pine Stands of the Southeastern United States 1,2 3 1 4 DANIEL R. MILLER, CHRIS ASARO, CHRISTOPHER M. CROWE, AND DONALD A. DUERR J. Econ. Entomol. 104(4): 1245Ð1257 (2011); DOI: 10.1603/EC11051 ABSTRACT In 2006, we examined the ßight responses of 43 species of longhorn beetles (Coleoptera: Cerambycidae) to multiple-funnel traps baited with binary lure blends of 1) ipsenol ϩ ipsdienol, 2) ethanol ϩ ␣-pinene, and a quaternary lure blend of 3) ipsenol ϩ ipsdienol ϩ ethanol ϩ ␣-pinene in the southeastern United States. In addition, we monitored responses of Buprestidae, Elateridae, and Curculionidae commonly associated with pine longhorn beetles. Field trials were conducted in mature pine (Pinus pp.) stands in Florida, Georgia, Louisiana, and Virginia. The following species preferred traps baited with the quaternary blend over those baited with ethanol ϩ ␣-pinene: Acanthocinus nodosus (F.), Acanthocinus obsoletus (Olivier), Astylopsis arcuata (LeConte), Astylopsis sexguttata (Say), Monochamus scutellatus (Say), Monochamus titillator (F.) complex, Rhagium inquisitor (L.) (Cerambycidae), Buprestis consularis Gory, Buprestis lineata F. (Buprestidae), Ips avulsus (Eichhoff), Ips calligraphus (Germar), Ips grandicollis (Eichhoff), Orthotomicus caelatus (Eichhoff), and Gna- thotrichus materiarus (Fitch) (Curculionidae). The addition of ipsenol and ipsdienol had no effect on catches of 17 other species of bark and wood boring beetles in traps baited with ethanol and ␣-pinene. Ethanol ϩ ␣-pinene interrupted the attraction of Ips avulsus, I. grandicollis, and Pityophthorus Eichhoff spp. (but not I. calligraphus) (Curculionidae) to traps baited with ipsenol ϩ ipsdienol. Our results support the use of traps baited with a quaternary blend of ipsenol ϩ ipsdienol ϩ ethanol ϩ ␣-pinene for common saproxylic beetles in pine forests of the southeastern United States. -

New Longhorn Beetles (Coleoptera: Cerambycidae) from Serbia
Arch. Biol. Sci., Belgrade, 57 (4), 27P-28P, 2005. NEW LONGHORN BEETLES (COLEOPTERA: CERAMBYCIDAE) FROM SERBIA. Nataša Pil1 and D. Stojanović2. 1Institute for Nature Conservation of Serbia, 21000 Novi Sad, Serbia and Montenegro, 2”Fruška Gora” National Park, 21208 Sremska Kamenica, Serbia and Montenegro UDC 597.76(497.11) Since the 1980’s, longhorn beetles (Coleoptera, Cerambycidae) They feed in the central region of the cone or occasionally in have been only randomly researched in Serbia. From earlier the base of old scales. The life cycle probably last two years, years, there are very detailed publications on this insect group and pupation very likely occurs in the soil. Adults emerge in (A d a m o v i ć , 1965; M i k š i ć and G e o r g i j e v i ć , 1971; April-July, on flowers. The given species differs from the simi- 1973; M i k š i ć and K o r p i č , 1985). lar Cortodera humeralis (Schaller, 1783) in having only sparse pubescence on the pronotum and head, with glabrous median The most recent data (I l i ć , 2005) indicate the presence line, and sparse pubescence on the outer border of the eye and of 245 longhorn beetle species (Coleoptera: Cerambycidae) in base of the antennae. Serbia. Not included in the mentioned publication, the follow- ing five species should be added to the list: Cortodera discolor 3. Vadonia hirsuta (Daniel and Daniel,1891) Fairmaire, 1866; Stenopterus similatus Holzschuh, 1979; Chlo- rophorus aegyptiacus (Fabricius, 1775); Agapanthia osmanlis (New data: Mt. -

Coleoptera, Cerambycidae, Lamiinae) with Description of a Species with Non‑Retractile Parameres
ARTICLE Two new genera of Desmiphorini (Coleoptera, Cerambycidae, Lamiinae) with description of a species with non‑retractile parameres Francisco Eriberto de Lima Nascimento¹² & Antonio Santos-Silva¹³ ¹ Universidade de São Paulo (USP), Museu de Zoologia (MZUSP). São Paulo, SP, Brasil. ² ORCID: http://orcid.org/0000-0002-5047-8921. E-mail: [email protected] ³ ORCID: http://orcid.org/0000-0001-7128-1418. E-mail: [email protected] Abstract. In this study, two new genera of Desmiphorini (Lamiinae) are proposed: Cleidaria gen. nov., to include Cleidaria cleidae sp. nov. from the state of Chiapas in Mexico, and Obscenoides gen. nov. for Desmiphora (D.) compta Martins & Galileo, 2005. The shape of tarsal claws of Cleidaria cleidae sp. nov. (abruptly narrowed from basal half) is so far, not found in any current genera of the tribe. With respect to Obscenoides compta (Martins & Galileo, 2005) comb. nov., the genitalia of males have an unusual shape with non-retractile parameres. The character combination related to this genital structure is unknown to us in other species in the family, and hypotheses about its function are suggested. Key-Words. Genital morphology; Longhorned beetles; New taxa; Taxonomy. INTRODUCTION the current definitions of some tribes, especially based on the works of Breuning do not take into Lamiinae (Cerambycidae), also known as flat- account adaptive convergences and use superfi- faced longhorns, with more than 21,000 described cial characters to subordinate taxa. species in about 3,000 genera and 87 tribes is Among these tribes, Desmiphorini Thomson, the largest subfamily of Cerambycidae occurring 1860 is not an exception, and its “boundaries” are worldwide (Tavakilian & Chevillotte, 2019). -

Analysis of the Breathing Apparatus of the Cerambycid Species That
UNIVERSIDADE DE LISBOA FACULDADE DE CIÊNCIAS DEPARTAMENTO DE BIOLOGIA ANIMAL Analysis of the breathing apparatus of the cerambycid species that colonizes pines infected by the pine wood nematode, Bursaphelenchus xylophilus (Steiner and Buhrer, 1934) Nickle, 1970, with special emphasis to the insect-vector Monochamus galloprovincialis (Olivier, 1795). Ana Catarina Guerreiro Leal Mestrado em Ecologia e Gestão Ambiental Dissertação orientada por: Professora Maria Teresa Rebelo (FCUL) Professor Manuel Francisco Pereira (IST) 2020 Agradecimentos À Professora Teresa Rebelo, que durante as suas aulas de Gestão Integrada de Pragas despertou o meu interesse e curiosidade nesta área e que aceitou comigo este desafio. Obrigada por toda a orientação, disponibilidade, conselho, paciência e encorajamento. Apesar da sua agenda conseguia sempre receber- me, ouvir as minha dúvidas e inquietações e mostrar-me um caminho para prosseguir com o trabalho. Ao Professor Manuel Francisco Pereira, que apesar da sua área de formação e trabalho não ser virada para a ‘bicharada’, foi um curioso e também aceitou este desafio. Obrigada por me receber no IST, pelas horas e horas de aquisição e pela disponibilidade em ensinar à bióloga ‘umas coisinhas’ sobre este ‘mundo’ que é a Micro-CT. Não posso não agradecer pelos bolinhos de canela que tinha no laboratório e que foram uma ajuda durante as nossas horas de análise de imagens. Ao Dr. Luís, pelo contagiante entusiasmo que sempre demonstra, e que me incentivou a oferecer- me para fazer parte deste projeto. Agradeço por todo o apoio no desenho experimental, por todo o material bibliográfico e biológico, pela disponibilidade em ouvir as minhas preocupações. À Professora Ana Cristina Figueiredo, por partilhar comigo o seu método de preparação de amostras para observação em SEM, pelos seus grandes contributos para agilizar a fixação e a liofilização dos insetos, por ter a paciência de repetir o protocolo até eu o dominar sozinha e por me abrir as portas do laboratório e me deixar à vontade. -

25Th U.S. Department of Agriculture Interagency Research Forum On
US Department of Agriculture Forest FHTET- 2014-01 Service December 2014 On the cover Vincent D’Amico for providing the cover artwork, “…and uphill both ways” CAUTION: PESTICIDES Pesticide Precautionary Statement This publication reports research involving pesticides. It does not contain recommendations for their use, nor does it imply that the uses discussed here have been registered. All uses of pesticides must be registered by appropriate State and/or Federal agencies before they can be recommended. CAUTION: Pesticides can be injurious to humans, domestic animals, desirable plants, and fish or other wildlife--if they are not handled or applied properly. Use all pesticides selectively and carefully. Follow recommended practices for the disposal of surplus pesticides and pesticide containers. Product Disclaimer Reference herein to any specific commercial products, processes, or service by trade name, trademark, manufacturer, or otherwise does not constitute or imply its endorsement, recom- mendation, or favoring by the United States government. The views and opinions of wuthors expressed herein do not necessarily reflect those of the United States government, and shall not be used for advertising or product endorsement purposes. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, sex, religion, age, disability, political beliefs, sexual orientation, or marital or family status. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at 202-720-2600 (voice and TDD). To file a complaint of discrimination, write USDA, Director, Office of Civil Rights, Room 326-W, Whitten Building, 1400 Independence Avenue, SW, Washington, D.C. -
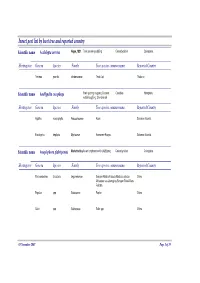
Insect Pest List by Host Tree and Reported Country
Insect pest list by host tree and reported country Scientific name Acalolepta cervina Hope, 1831 Teak canker grub|Eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Tectona grandis Verbenaceae Teak-Jati Thailand Scientific name Amblypelta cocophaga Fruit spotting bug|eng Coconut Coreidae Hemiptera nutfall bug|Eng, Chinche del Hosting tree Genera Species Family Tree species common name Reported Country Agathis macrophylla Araucariaceae Kauri Solomon Islands Eucalyptus deglupta Myrtaceae Kamarere-Bagras Solomon Islands Scientific name Anoplophora glabripennis Motschulsky Asian longhorn beetle (ALB)|eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Paraserianthes falcataria Leguminosae Sengon-Albizia-Falcata-Molucca albizia- China Moluccac sau-Jeungjing-Sengon-Batai-Mara- Falcata Populus spp. Salicaceae Poplar China Salix spp. Salicaceae Salix spp. China 05 November 2007 Page 1 of 35 Scientific name Aonidiella orientalis Newstead, Oriental scale|eng Diaspididae Homoptera 1894 Hosting tree Genera Species Family Tree species common name Reported Country Lovoa swynnertonii Meliaceae East African walnut Cameroon Azadirachta indica Meliaceae Melia indica-Neem Nigeria Scientific name Apethymus abdominalis Lepeletier, Tenthredinidae Hymenoptera 1823 Hosting tree Genera Species Family Tree species common name Reported Country Other Coniferous Other Coniferous Romania Scientific name Apriona germari Hope 1831 Long-horned beetle|eng Cerambycidae