Assessing the Most Irreplaceable Protected Areas for the Conservation of Mammals in the Atlantic Forest: Lessons for the Governance of Mosaics
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
314 Trumbull Was Apparently Ill-Advised Again, Possibly By
314 BOOK REVIEWS Trumbull was apparently ill-advised again, possibly by expatriate administrators, as seen in his claim that, , Finally, the leadership class already existing under ancient custon is being expanded in depth everywhere by the spread of education, and I have found no significant disposition among the conservative oldsters to stifle the progressive young, despite the generation gap that obtrudes in family life. (282) Most observers of Micronesia would agree that one of the saddest aspects of develop- ment and change in the U.S. administered islands is the fact that the traditional leaders were, for the most part, by-passed in both education and political development. Of all the traditional leaders of Micronesia, there are but a few who are reasonably conver- sant in English, which is the language of government. None of the traditional para- mount chiefs holds an elected office of any consequence. Since I seem to be picking at the author, I may as well advise him to avoid giving spurious etymologies of exotic words. The item he cites as ghoose(p. 135) doesn't mean "bribe" or anything else in any of the languages of Fiji. Nor does Guahan by itself mean "we have". It is simply a place name, the Chamorro version of Guam. Possibly the best feature of this book is the anecdotal and personal touches that come from Mr. Trumbull's familiarity with and love for the islands themselves. Like many a Pacific War vet, he feels for the islands in a very special way which comes through in his writing. -

Hawker Policy in Thailand
Legislative Council Secretariat FS12/13-14 FACT SHEET Hawker policy in Thailand 1. Background 1.1 Similar to many other Asian countries, Thailand has a long history of street vending. In Bangkok, the largest city in Thailand, street vending has provided local Thai people with cheap and convenient access to a wide range of goods and a means of making a living. According to a survey conducted by the International Labour Office1, a majority of street vendors surveyed were satisfied with their occupation because of the income earning opportunity and work autonomy.2 In recent years, street vending has also been considered as a way to nurture entrepreneurship, as well as adding to the tourist attractions of Bangkok by bringing vibrancy and vitality to the city. 1.2 In Bangkok, street vending has brought with it urban problems such as obstruction to pedestrian and vehicular traffic. It has also given rise to hygienic problem as many of street vending activities are related to the sale of cooked food along the streets. As such, the local government has designated various locations as street vending areas subject to regulatory controls such as the restrictions on the trading hours of street vendors. This fact sheet makes reference to the Bangkok city for the study of hawker policy in Thailand, covering information on the regulation and management of street vending, different forms of government-run markets, and the emergence of the new-generation street vendors. 1 International Labour Office is the permanent secretariat of the International Labour Organization. 2 See International Labour Office (2006a). -

Top of Page Interview Information--Different Title
Oral History Center University of California The Bancroft Library Berkeley, California The Freedom to Marry Oral History Project Thalia Zepatos Thalia Zepatos on Research and Messaging in Freedom to Marry Interviews conducted by Martin Meeker in 2016 Copyright © 2017 by The Regents of the University of California ii Since 1954 the Oral History Center of the Bancroft Library, formerly the Regional Oral History Office, has been interviewing leading participants in or well-placed witnesses to major events in the development of Northern California, the West, and the nation. Oral History is a method of collecting historical information through tape-recorded interviews between a narrator with firsthand knowledge of historically significant events and a well-informed interviewer, with the goal of preserving substantive additions to the historical record. The tape recording is transcribed, lightly edited for continuity and clarity, and reviewed by the interviewee. The corrected manuscript is bound with photographs and illustrative materials and placed in The Bancroft Library at the University of California, Berkeley, and in other research collections for scholarly use. Because it is primary material, oral history is not intended to present the final, verified, or complete narrative of events. It is a spoken account, offered by the interviewee in response to questioning, and as such it is reflective, partisan, deeply involved, and irreplaceable. ********************************* All uses of this manuscript are covered by a legal agreement between The Regents of the University of California and Thalia Zepatos dated September 6, 2016. The manuscript is thereby made available for research purposes. All literary rights in the manuscript, including the right to publish, are reserved to The Bancroft Library of the University of California, Berkeley. -

Remuneração Dos Vereadores E Jornada De Trabalho
REMUNERAÇÃO DOS VEREADORES E JORNADA DE TRABALHO François E J. de Bremaeker Salvador, março de 2010 2 Estão sendo apresentados por Município os dados levantados pelo Interlegis – Comunidade Virtual do Poder Legislativo – do Senado Federal, sobre a remuneração dos Vereadores em 2005, sua conversão em salários mínimos, a projeção do valor para 2010, relacionado ao IPCA e ao salário mínimo e, a jornada de trabalho. Valores “zero” e “não respondeu” correspondem à “não informado”. REMUN SALAR ESTIM ESTIM JORNADA MUNICÍPIOS 2005 MINIM IPCA SAL MIN DE 2005 2010 2010 TRABALHO RONDÔNIA ALTA FLORESTA DO OESTE 2.500 8,33 3.145 4.250 12 HORAS ALTO ALEGRE DOS PARECIS 1.600 5,33 2.012 2.720 8 HORAS ALTO PARAISO 1.550 5,17 1.950 2.635 8 HORAS ALVORADA DO OESTE 0 0,00 0 0 NÃO RESPONDEU ARIQUEMES 4.400 14,67 5.534 7.480 20 HORAS BURITIS 0 0,00 0 0 NÃO RESPONDEU CABIXI 680 2,27 855 1.156 ACIMA DE 40 HORAS CACAULANDIA 1.100 3,67 1.384 1.870 12 HORAS CACOAL 3.600 12,00 4.528 6.120 20 HORAS CAMPO NOVO DE RONDONIA 0 0,00 0 0 NÃO RESPONDEU CANDEIAS DO JAMARI 2.000 6,67 2.516 3.400 8 HORAS CASTANHEIRAS 800 2,67 1.006 1.360 8 HORAS CEREJEIRAS 1.745 5,82 2.195 2.967 24 HORAS CHUPINGUAIA 1.900 6,33 2.390 3.230 8 HORAS COLORADO DO OESTE 1.564 5,21 1.967 2.659 8 HORAS CORUMBIARA 900 3,00 1.132 1.530 8 HORAS COSTA MARQUES 0 0,00 0 0 NÃO RESPONDEU CUJUBIM 1.118 3,73 1.406 1.901 20 HORAS ESPIGAO DOESTE 1.800 6,00 2.264 3.060 12 HORAS GOVERNADOR JORGE TEIXEIRA 1.912 6,37 2.405 3.250 16 HORAS GUAJARA MIRIM 2.115 7,05 2.660 3.596 12 HORAS ITAPUA DO OESTE 750 2,50 943 1.275 -

Histosols in an Upper Montane Environment in the Itatiaia Plateau
Rev Bras Cienc Solo 2016;40:e0160176 Article Division – Soil in Space and Time | Commission – Soil Survey and Classification Histosols in an Upper Montane Environment in the Itatiaia Plateau Paula Fernanda Chaves Soares(1), Lúcia Helena Cunha dos Anjos(2), Marcos Gervasio Pereira(2)* and Luiz Carlos Ruiz Pessenda(3) (1) Universidade Federal Rural do Rio de Janeiro, Instituto de Agronomia, Departamento de Solos, Programa de Pós-graduação em Agronomia – Ciência do Solo, Seropédica, Rio de Janeiro, Brasil. (2) Universidade Federal Rural do Rio de Janeiro, Instituto de Agronomia, Departamento de Solos, Seropédica, Rio de Janeiro, Brasil. (3) Universidade de São Paulo, Escola de Agricultura Luiz de Queiroz, Centro de Energia Nuclear na Agricultura, Piracicaba, São Paulo, Brasil. ABSTRACT: Highland environments favor accumulation and preservation of soil organic matter (SOM) due to low temperatures, leading to the formation of Histosols. The Itatiaia National Park (INP), Rio de Janeiro, Brazil, offers conditions for preservation of SOM deposited over time, which has led to the formation of these soils. The objective of this study was to characterize Histosols in this environment, with the premise that it may provide evidence of changes in vegetation. Organossolos (Histosols) were sampled, characterized, described, and analyzed for their properties, stable isotopes of 12C and 13C, 14N and 15N, and dating of organic matter through 14C. The soils were classified in the Brazilian Soil Classification System as Organossolo Háplico Hêmico típico - RJ-01 (Haplohemists) and Organossolo Fólico Sáprico cambissólico - RJ-02 (Udifolists). The morphological properties, degree of transformation, and chemical fractioning of SOM were consistent with hemic and sapric materials. -

Dalechampia COM ÊNFASE EM Dalechampia Sect
RAFAELA ALVES PEREIRA DA SILVA FILOGENIA E TAXONOMIA DE Dalechampia COM ÊNFASE EM Dalechampia sect. Dalechampia, Euphorbiaceae RECIFE 2019 I RAFAELA ALVES PEREIRA DA SILVA FILOGENIA E TAXONOMIA DE Dalechampia COM ÊNFASE EM Dalechampia sect. Dalechampia, Euphorbiaceae Tese apresentada ao Programa de Pós-Graduação em Botânica da Universidade Federal Rural de Pernambuco, como parte dos requisitos para obtenção do título de Doutora em Botânica. Orientadora: Profª. Dra. Margareth Ferreira de Sales Deptº de Biologia, Área de Botânica/UFRPE Co-orientadores: Dra. Sarah Maria Athiê-Souza Profº. Dr. Luís Gustavo Rodrigues de Souza Colaborador: Dr. Scott Armbruster Profº. Dr. André Laurênio de Melo RECIFE 2019 II Dados Internacionais de Catalogação na Publicação (CIP) Sistema Integrado de Bibliotecas da UFRPE Biblioteca Central, Recife-PE, Brasil S586f Silva, Rafaela Alves Pereira da. Filogenia e taxonomia de Dalechampia com ênfase em Dalechampia sect. Dalechampia, Euphorbiaceae / Rafaela Alves Pereira da Silva. – Recife, 2019. 335 f.: il. Orientador(a): Margareth Ferreira de Sales. Coorientador(a): Sarah Maria Athiê-Souza Coorientador(a): Luís Gustavo Rodrigues de Souza Tese (Doutorado) – Universidade Federal Rural de Pernambuco, Programa de Pós-Graduação em Botânica, Recife, BR-PE, 2019. Inclui referências e apêndice(s). 1. Dalechampiinae 2. Biogeography 3. Molecular 4. Character evolution I. Sales, Margareth Ferreira de, orient. II. Athiê-Souza, Sarah Maria, coorient. III Souza, Luís Gustavo Rodrigues de, coorient. IV. Título CDD 581 III FILOGENIA E TAXONOMIA DE Dalechampia COM ÊNFASE EM Dalechampia sect. Dalechampia, Euphorbiaceae IV Dedico Ao Espírito Santo de Deus. Ofereço A Ednaldo José da Silva “Ignore aquele que diz: você não tem valor por isso ou por aquilo, porque eu te amo muito e torço por você”. -

Distribution of Birds Along an Elevational Gradient in the Atlantic Forest of Brazil: Implications for the Conservation of Endemic and Endangered Species
Bird Conservation International (1999) 9:235-253. © BirdLife International 1999 Distribution of birds along an elevational gradient in the Atlantic forest of Brazil: implications for the conservation of endemic and endangered species JAQUELINE M. GOERCK Summary In this study I compare bird communities along an elevational gradient in an Atlantic forest remnant (Pico do Corcovado in Ubatuba) in coastal Sao Paulo state, Brazil. Forests at low elevations are structurally more complex and more diverse in plant species than those along the slopes and at higher elevations in this remnant. Consequently it is hypothesized that low elevation forests contain a greater diversity of bird species. Results from the study in the Corcovado area show clear differences in the distribution of forest birds along the elevational gradient from both qualitative and quantitative aspects. The structurally more complex forest at low elevations contains the most diverse avifauna, including several of the rarest and most threatened species. The importance of this remnant as a whole is apparent due to the high diversity observed (254 species), the high proportion of endemic species, and the extent to which the avifauna is endangered. Protection of forests at all elevations along the Serra do Mar is required to maintain diversity of bird species, particularly the many endemic and endangered species restricted to specific elevations. Resumo Comunidades de aves foram comparadas ao longo de um gradiente altitudinal em um remanescente florestal de Mata Atlantica (Pico do Corcovado em Ubatuba) na costa do estado de Sao Paulo. As florestas de baixada sao mais diversas e mais complexas estruturalmente do que as florestas de encosta ou de altitude e consequentemente espera-se que as florestas de baixada possuam uma maior diversidade de aves. -

Mamíferos Não Voadores Do Parque Estadual Carlos Botelho, Continuum Florestal Do Paranapiacaba
Biota Neotrop., vol. 12, no. 4 Mamíferos não voadores do Parque Estadual Carlos Botelho, Continuum florestal do Paranapiacaba Carlos Rodrigo Brocardo1,2,3, Raisa Rodarte1, Rafael da Silveira Bueno1, Laurence Culot1 & Mauro Galetti1 1Laboratório de Biologia da Conservação, Departamento de Ecologia, Universidade Estadual Paulista – UNESP, CP 199, CEP 13506-900, Rio Claro, SP, Brasil 2Instituto Neotropical: Pesquisa e Conservação, CP 19009, CEP 81531-980, Curitiba, PR, Brasil 3Autor para correspondência: Carlos Rodrigo Brocardo, e-mail: [email protected] BROCARDO, C.R., RODARTE, R., BUENO, R.S., CULOT, L. & GALETTI, M. Non-volant mammals of Carlos Botelho State Park, Paranapiacaba Forest Continuum. Biota Neotrop. 12(4): http://www.biotaneotropica.org. br/v12n4/pt/abstract?inventory+bn02512042012 Abstract: The Atlantic Forest is one of the most studied Brazilian biomes in relation to its mammalian fauna. However, there is still a series of gaps of knowledge about the distribution and persistence of some of these species which prevents taking adequate conservation measures to better protect the mammals. In order to make the inventory of the non-volant mammalian fauna of the Carlos Botelho State Park (SP), we compiled data of camera trapping, diurnal census, track records, pitfall and live trapping collected over 8 years (2004-2012). We registered a total of 53 species, of which 12 are regionally threatened and one is an exotic species (Lepus europaeus), including the presence of most mammal species expected for the Paranapiacaba Forest Continuum. The high non-volant mammals species richness allied to the presence of threatened species, strengthen the role of this protected area for mammal conservation in the Atlantic Forest. -

The Globalization of Chinese Food ANTHROPOLOGY of ASIA SERIES Series Editor: Grant Evans, University Ofhong Kong
The Globalization of Chinese Food ANTHROPOLOGY OF ASIA SERIES Series Editor: Grant Evans, University ofHong Kong Asia today is one ofthe most dynamic regions ofthe world. The previously predominant image of 'timeless peasants' has given way to the image of fast-paced business people, mass consumerism and high-rise urban conglomerations. Yet much discourse remains entrenched in the polarities of 'East vs. West', 'Tradition vs. Change'. This series hopes to provide a forum for anthropological studies which break with such polarities. It will publish titles dealing with cosmopolitanism, cultural identity, representa tions, arts and performance. The complexities of urban Asia, its elites, its political rituals, and its families will also be explored. Dangerous Blood, Refined Souls Death Rituals among the Chinese in Singapore Tong Chee Kiong Folk Art Potters ofJapan Beyond an Anthropology of Aesthetics Brian Moeran Hong Kong The Anthropology of a Chinese Metropolis Edited by Grant Evans and Maria Tam Anthropology and Colonialism in Asia and Oceania Jan van Bremen and Akitoshi Shimizu Japanese Bosses, Chinese Workers Power and Control in a Hong Kong Megastore WOng Heung wah The Legend ofthe Golden Boat Regulation, Trade and Traders in the Borderlands of Laos, Thailand, China and Burma Andrew walker Cultural Crisis and Social Memory Politics of the Past in the Thai World Edited by Shigeharu Tanabe and Charles R Keyes The Globalization of Chinese Food Edited by David Y. H. Wu and Sidney C. H. Cheung The Globalization of Chinese Food Edited by David Y. H. Wu and Sidney C. H. Cheung UNIVERSITY OF HAWAI'I PRESS HONOLULU Editorial Matter © 2002 David Y. -

Parques Nacionais
National Parks Brazil BrasiParques Nacionails Brasil Parques Nacionais 2 3 4 5 National Parks Brazil BrasiParques Nacionails 6 7 O Brasil em sua imensidão abriga hoje 69 parques nacionais Brazil in its immensity today houses 69 national parks located situados nas cinco macro-regiões, protegendo no Norte áreas de in the five macro-regions, protecting the northern areas of florestas virgens e praticamente intocadas pelo homem, dunas e virgin forests – virtually untouched by man, dunes and rock pinturas rupestres no Nordeste, a exuberância de Mata Atlântica paintings in the Northeast, the exuberance of the Southeast no Sudeste, os Campos Gerais no Sul e uma flora e fauna do Atlantic Forest, Campos Gerais in the South and the exuberant exuberante do Cerrado no Centro-Oeste. Através desta publica- flora and fauna of the Cerrado in the Midwest. Through this ção a Localiza disponibiliza mais uma vez aos seus clientes e publication, Localiza makes available once more to its clients leitores a possibilidade de descoberta de exemplos bem suce- and readers the chance of discovering successful examples didos de manutenção da riqueza natural, legando às próximas of the maintenance of natural wealth, bequeathing to future gerações áreas de rara beleza. Juntas, elas compõem hoje um generations areas of outstanding beauty. Together they rico mosaico de preservação de nossa inigualável biodiversida- compose today a rich mosaic of conservation of our unique de, de nossa história e também nossa cultura. biodiversity, our history and our culture. Apoio Patrocínio Realização 8 9 Em 1876 o engenheiro abolicionista negro André Rebouças, foi precursor ao idealizar que o Brasil In 1876, the abolitionist engineer André Rebouças was a precursor when he idealized that Brazil destinasse parte de seu território para a criação de áreas protegidas com o intuito de salvaguardar would separate part of its territory to create protected areas with the intention to safeguard in a de forma sistemática, legal e organizada, aspectos importantes de nossos ecossistemas regionais. -
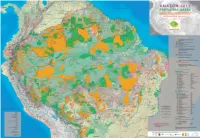
Mapaamazonia2012-Deforestationing
Amazon and human population Bolivia Brasil Colombia Ecuador Guyana Guyane Française Perú Suriname Venezuela total Amazon Protected Areas and Indigenous Territories % of the national % of the national % of the national % of the national % of the national % of the national % of the national % of the national % of the national % of the Amazon total total total total total total total total total total In Amazon the protection of socioenvironmental diversity is being consolidated through the recognition of the territorial rights of indigenous peoples A M A Z O N 2012 Total population and the constitution of a varied set of protected areas. These conservation strategies have been expanding over recent years and today cover a 8,274,325 - 191,480,630 - 42,090,502 - 14,483,499 - 751,000 - 208,171 - 28,220,764 - 492,829 - 27,150,095 - 313,151,815 (nº of inhabitants) surface area of 3,502,750 km2 – 2,144,412 km2 in Indigenous Territories and 1,696,529 km2 in Protected Natural Areas, with an overlap of 336,365 2 Amazon population km between them – which corresponds to 45% of the region. PROTECTED AREAS and INDIGENOUS TERRITORIES 1,233,727 14.9% 23,654,336 12.4% 1,210,549 2.9% 739,814 5.1% 751,000 100.0% 208,171 100.0% 3,675,292 13.0% 492,829 100.0% 1,716,984 6.3% 33,682,702 10.8% (nº of inhabitants) The challenge faced in terms of attaining the objectives of strengthening the cultural and biological diversity of Amazon, represented in indigenous Total area of the country (km2) 1,098,581 - 8,514,876 - 1,141,748 - 249,041 - 214,969 - 86,504 - 1,285,215 - 163,820 - 916,445 - 13,671,199 territories and protected areas, encompasses a variety of aspects. -

Large and Medium-Sized Mammals of Nova Baden State Park, Minas Gerais, Brazil
13 3 the journal of 2141 biodiversity data 13 June 2017 Check List LISTS OF SPECIES Check List 13(3): 2141, 13 June 2017 https://doi.org/10.15560/13.3.2141 ISSN 1809-127X © 2017 Check List and Authors Large and medium-sized mammals of Nova Baden State Park, Minas Gerais, Brazil Clarissa Alves da Rosa1, 2 & Agnis Cristiane Souza1 1 Independent researcher, Street 3600, n 232, 88330 248, Balneário Camboriú, SC, Brazil 2 Corresponding author. E-mail: [email protected] Abstract. Nova Baden State Park (NBSP) is located in the gaps in scientific information about their biodiversity, espe- Brazilian Atlantic Forest which is a biodiversity hotspot and cially large and medium-sized mammals. Thus our aim is to priority for conservation. Our aim is to provide a list of large provide a list of medium and large-sized mammals in Nova and medium-sized mammal species recorded in NBSP. We Baden State Park (NBSP), a protected area in the Serra da made a camera trap survey and opportunistic observations Mantiqueira, Minas Gerais state. from December 2014 to September 2015, and searched the grey literature. We recorded 12 large and medium-sized mammals in MATERIALS AND METHODS our survey and 11 more species listed in grey literature. The 23 species registered for NBSP belong to eight orders (Carnivora, Study site Primates, Rodents, Cingulata, Pilosa, Didephimorphia, NBSP is located in southeastern region of Brazil at munici- Lagomorpha and Artiodactyla), including threatened species at pally of Lambari, Minas Gerais (21°56ʹ00ʺ S, 045°18ʹ30ʺ W, local, national and global levels. With an important mammal datum WGS84).