Alien Vascular Plants Recorded from the Barentsburg and Pyramiden Settlements, Svalbard
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Handbok07.Pdf
- . - - - . -. � ..;/, AGE MILL.YEAR$ ;YE basalt �- OUATERNARY votcanoes CENOZOIC \....t TERTIARY ·· basalt/// 65 CRETACEOUS -� 145 MESOZOIC JURASSIC " 210 � TRIAS SIC 245 " PERMIAN 290 CARBONIFEROUS /I/ Å 360 \....t DEVONIAN � PALEOZOIC � 410 SILURIAN 440 /I/ ranite � ORDOVICIAN T 510 z CAM BRIAN � w :::;: 570 w UPPER (J) PROTEROZOIC � c( " 1000 Ill /// PRECAMBRIAN MIDDLE AND LOWER PROTEROZOIC I /// 2500 ARCHEAN /(/folding \....tfaulting x metamorphism '- subduction POLARHÅNDBOK NO. 7 AUDUN HJELLE GEOLOGY.OF SVALBARD OSLO 1993 Photographs contributed by the following: Dallmann, Winfried: Figs. 12, 21, 24, 25, 31, 33, 35, 48 Heintz, Natascha: Figs. 15, 59 Hisdal, Vidar: Figs. 40, 42, 47, 49 Hjelle, Audun: Figs. 3, 10, 11, 18 , 23, 28, 29, 30, 32, 36, 43, 45, 46, 50, 51, 52, 53, 54, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 71, 72, 75 Larsen, Geir B.: Fig. 70 Lytskjold, Bjørn: Fig. 38 Nøttvedt, Arvid: Fig. 34 Paleontologisk Museum, Oslo: Figs. 5, 9 Salvigsen, Otto: Figs. 13, 59 Skogen, Erik: Fig. 39 Store Norske Spitsbergen Kulkompani (SNSK): Fig. 26 © Norsk Polarinstitutt, Middelthuns gate 29, 0301 Oslo English translation: Richard Binns Editor of text and illustrations: Annemor Brekke Graphic design: Vidar Grimshei Omslagsfoto: Erik Skogen Graphic production: Grimshei Grafiske, Lørenskog ISBN 82-7666-057-6 Printed September 1993 CONTENTS PREFACE ............................................6 The Kongsfjorden area ....... ..........97 Smeerenburgfjorden - Magdalene- INTRODUCTION ..... .. .... ....... ........ ....6 fjorden - Liefdefjorden................ 109 Woodfjorden - Bockfjorden........ 116 THE GEOLOGICAL EXPLORATION OF SVALBARD .... ........... ....... .......... ..9 NORTHEASTERN SPITSBERGEN AND NORDAUSTLANDET ........... 123 SVALBARD, PART OF THE Ny Friesland and Olav V Land .. .123 NORTHERN POLAR REGION ...... ... 11 Nordaustlandet and the neigh- bouring islands........................... 126 WHA T TOOK PLACE IN SVALBARD - WHEN? .... -

Climate in Svalbard 2100
M-1242 | 2018 Climate in Svalbard 2100 – a knowledge base for climate adaptation NCCS report no. 1/2019 Photo: Ketil Isaksen, MET Norway Editors I.Hanssen-Bauer, E.J.Førland, H.Hisdal, S.Mayer, A.B.Sandø, A.Sorteberg CLIMATE IN SVALBARD 2100 CLIMATE IN SVALBARD 2100 Commissioned by Title: Date Climate in Svalbard 2100 January 2019 – a knowledge base for climate adaptation ISSN nr. Rapport nr. 2387-3027 1/2019 Authors Classification Editors: I.Hanssen-Bauer1,12, E.J.Førland1,12, H.Hisdal2,12, Free S.Mayer3,12,13, A.B.Sandø5,13, A.Sorteberg4,13 Clients Authors: M.Adakudlu3,13, J.Andresen2, J.Bakke4,13, S.Beldring2,12, R.Benestad1, W. Bilt4,13, J.Bogen2, C.Borstad6, Norwegian Environment Agency (Miljødirektoratet) K.Breili9, Ø.Breivik1,4, K.Y.Børsheim5,13, H.H.Christiansen6, A.Dobler1, R.Engeset2, R.Frauenfelder7, S.Gerland10, H.M.Gjelten1, J.Gundersen2, K.Isaksen1,12, C.Jaedicke7, H.Kierulf9, J.Kohler10, H.Li2,12, J.Lutz1,12, K.Melvold2,12, Client’s reference 1,12 4,6 2,12 5,8,13 A.Mezghani , F.Nilsen , I.B.Nilsen , J.E.Ø.Nilsen , http://www.miljodirektoratet.no/M1242 O. Pavlova10, O.Ravndal9, B.Risebrobakken3,13, T.Saloranta2, S.Sandven6,8,13, T.V.Schuler6,11, M.J.R.Simpson9, M.Skogen5,13, L.H.Smedsrud4,6,13, M.Sund2, D. Vikhamar-Schuler1,2,12, S.Westermann11, W.K.Wong2,12 Affiliations: See Acknowledgements! Abstract The Norwegian Centre for Climate Services (NCCS) is collaboration between the Norwegian Meteorological In- This report was commissioned by the Norwegian Environment Agency in order to provide basic information for use stitute, the Norwegian Water Resources and Energy Directorate, Norwegian Research Centre and the Bjerknes in climate change adaptation in Svalbard. -

S V a L B a R D Med Fastboende I Ny-Ålesund
Kart B i forskrift om motorferdsel på Svalbard Map B in regulations realting to motor traffic in Svalbard Ferdsel med beltemotorsykkel (snøskuter) og beltebil på Svalbard - tilreisende jf. § 8 Område der tilreisende kan bruke beltemotorsykkel (snøskuter) og beltebil på snødekt og frossen mark. Område der tilreisende kan bruke beltemotorsykkel (snøskuter) og beltebil på snødekt og frossen mark dersom de deltar i organiserte turopplegg eller er i følge med fastboende. Ny-Ålesund! Område der tilreisende kan bruke beltemotorsykkel (snøskuter) og beltebil på snødekt og frossen mark dersom de er i følge S V A L B A R D med fastboende i Ny-Ålesund. Område for ikke-motorisert ferdsel. All snøskuterkjøring forbudt. Ferdselsåre der tilreisende kan bruke beltemotorsykkel (snøskuter) Pyramiden ! og beltebil på snødekt og frossen mark dersom de deltar i organiserte turopplegg eller er i følge med fastboende. På Storfjorden mellom Agardhbukta og Wichebukta skal ferdselen legges til nærmeste farbare vei på sjøisen langsetter land. Area where visitors may use snowmobiles and tracked vehicles in Svalbard, see section 8 of the regulations ! Longyearbyen Area where visitors may use snowmobiles and tracked vehicles on snow-covered and frozen ground. !Barentsburg Area where visitors may use snowmobiles and tracked vehicles on snow-covered and frozen ground if they are taking part in an organised tour or are accompanying permanent residents. Sveagruva ! Area where visitors may use snowmobiles and tracked vehicles on snow-covered and frozen ground if they are accompanying permanent residents of Ny-Ålesund. Area reserved for non-motor traffic. All snowmobile use is prohibited. Trail where visitors may use snowmobiles and tracked vehicles onsnow-covered and frozen ground if they are taking part in an organised tour or are accompanying permanent residents. -

Ny-Ålesund Research Station
Ny-Ålesund Research Station Research Strategy Applicable from 2019 DEL XX / SEKSJONSTITTEL Preface Svalbard research is characterised by a high degree of interna- tional collaboration. In Ny-Ålesund more than 20 research About the Research Council of Norway institutes have long-term research and monitoring activities. The station is one of four research localities in Svalbard (Ny-Ålesund, Longyearbyen, Barentsburg and Hornsund). The Research Council of Norway is a national strategic and research community, trade and industry and the public Close cooperation between these communities is essential funding agency for research activities. The Council serves as administration. It is the task of the Research Council to identify for the further development of Ny-Ålesund. the key advisor on research policy issues to the Norwegian Norway’s research needs and recommend national priorities Photo: John-Arne Røttingen Government, the government ministries, and other central and to use different funding schemes to help to translate In 2016, the Norwegian Government announced (Meld.St.32 institutions and groups involved in research and development national research policy goals into action. The Research Council (2015-2016)) the development of a research strategy for the (R&D). The Research Council also works to increase financial provides a central meeting place for those who fund, carry out Ny-Ålesund research station. Guidelines and principles for investment in, and raise the quality of, Norwegian R&D and and utilise research and works actively to promote the research activity were established by the government in 2018 to promote innovation in a collaborative effort between the internationalisation of Norwegian research. -

Svalbard (Norway)
Svalbard (Norway) Cross border travel - People - Depending on your citizenship, you may need a visa to enter Svalbard. - The Norwegian authorities do not require a special visa for entering Svalbard, but you may need a permit for entering mainland Norway /the Schengen Area, if you travel via Norway/the Schengen Area on your way to or from Svalbard. - It´s important to ensure that you get a double-entry visa to Norway so you can return to the Schengen Area (mainland Norway) after your stay in Svalbard! - More information can be found on the Norwegian directorate of immigration´s website: https://www.udi.no/en/ - Find more information about entering Svalbard on the website of the Governor of Svalbard: https://www.sysselmannen.no/en/visas-and-immigration/ - Note that a fee needs to be paid for all visa applications. Covid-19 You can find general information and links to relevant COVID-19 related information here: https://www.sysselmannen.no/en/corona-and-svalbard/ Note that any mandatory quarantine must be taken in mainland Norway, not on Svalbard! Find more information and quarantine (hotels) here: https://www.regjeringen.no/en/topics/koronavirus-covid- 19/the-corona-situation-more-information-about-quarantine- hotels/id2784377/?fbclid=IwAR0CA4Rm7edxNhpaksTgxqrAHVXyJcsDBEZrtbaB- t51JTss5wBVz_NUzoQ You can find further information regarding the temporary travel restrictions here: https://nyalesundresearch.no/covid-info/ - Instrumentation (import/export) - In general, it is recommended to use a shipping/transport agency. - Note that due to limited air cargo capacity to and from Ny-Ålesund, cargo related to research activity should preferably be sent by cargo ship. -
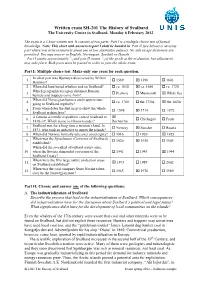
Written Exam SH-201 the History of Svalbard the University Centre in Svalbard, Monday 6 February 2012
Written exam SH-201 The History of Svalbard The University Centre in Svalbard, Monday 6 February 2012 The exam is a 3 hour written test. It consists of two parts: Part I is a multiple choice test of factual knowledge. Note: This sheet with answers to part I shall be handed in. Part II (see below) is an essay part where you write extensively about one of two alternative subjects. No aids except dictionary are permitted. You may answer in English, Norwegian, Swedish or Danish. 1 2 Part I counts approximately /3 and part II counts /3 of the grade at the evaluation, but adjustment may take place. Both parts must be passed in order to pass the whole exam. Part I: Multiple choice test. Make only one cross for each question. In what year was Bjørnøya discovered by Willem 1. 1569 1596 1603 Barentsz? 2. When did land-based whaling end on Svalbard? ca. 1630 ca. 1680 ca. 1720 Which geographical region did most Russian 3. Pechora Murmansk White Sea hunters and trappers come from? When did Norwegian hunters and trappers start 4. ca. 1700 the 1750s the 1820s going to Svalbard regularly? From when dates the first map to show the whole 5. 1598 1714 1872 Svalbard archipelago? A famous scientific expedition visited Svalbard in 6. Chichagov Fram 1838–39. Which name is it known under? Recherche Svalbard was for a long time a no man’s land. In 7. Norway Sweden Russia 1871, who took an initiative to annex the islands? 8. When did Norway formally take over sovereignty? 1916 1920 1925 When was the Sysselmann (Governor of Svalbard) 9. -

Arctic Territories Svalbard As a Fluid Territory Contents
ARCTIC TERRITORIES SVALBARD AS A FLUID TERRITORY CONTENTS Introduction ........................................................................... 01 Part I Study trip ................................................................................ 04 Site visits ................................................................................ 05 Fieldwork ............................................................................... 07 Identifying themes/subjects of interest ................................... 11 Introducing short sections ...................................................... 14 Part II Sections: Introduction ............................................................. 17 Sections: Finding a narrative .................................................. 19 Sections: Introducing time ...................................................... 20 Describing forces through glossaries ......................... 21 Describing forces through illustrations ....................... 22 Long sections .................................................................... 24 Part III Interaction Points: Introduction ............................................... 40 Interaction Points: Forces overlay .......................................... 41 Interaction points: Revealing archives .................................... 42 Model making - terrain model ................................................ 52 SVALBARD STUDIO - FALL SEMESTER 2015 MASTER OF LANDSCAPE ARCHITECTURE Part IV TROMSØ ACADEMY OF LANDSCAPE AND TERRITORIAL STUDIES Return to -

Telenor Ninety Years on Spitsbergen
Telenor ninety years on Spitsbergen This year, it will be ninety year since Telenor (at that time Telegrafvæsenet) established the first telephone link between Spitsbergen and the Norwegian mainland. With the exception of a few years during the second world war, Telenor has been present on Svalbard since 1911. This weekend, Telenor, one of Svalbard's pioneers, will be celebrating its continued active role in the island group through several arrangements. Spitsbergen Radio On May 3, 1911, the Norwegian parliament agreed that a radio telegraph station was to be built on Svalbard. The station was named Spitsbergen Radio and originally built at Green Harbour (now Grønfjorden). It was later moved to Finneset just south of Barentsburg. The station was established five years before Svalbard's first coal mine opened. The first telephone connection linking Svalbard to the mainland based on the technology of the time was made on November 22, 1911. A corresponding station was built on the mainland. In 1930, the station that had been named Svalbard Radio in 1925 was moved to Longyearbyen. Isfjord Radio In 1933, Isfjord Radio was established at Kapp Linne. The station was established to act as an intermediary for traffic between Svalbard Radio and ships in the waters around Svalbard. During the second world war, Isfjord Radio was decommissioned and destroyed by German occupying forces, but the station was rebuilt and set back into operation in 1946. Most of Isfjord Radio's operations were moved to Longyearbyen when the airport was opened in 1975. Today, Svalbard Radio operates from the airport's control tower, handling communication for both sea and air traffic. -

Winning Coal at 78° North : Mining, Contingency and the Chaîne Opératoire in Old Longyear City
Michigan Technological University Digital Commons @ Michigan Tech Dissertations, Master's Theses and Master's Dissertations, Master's Theses and Master's Reports - Open Reports 2009 Winning coal at 78° North : mining, contingency and the Chaîne Opératoire in old Longyear City Seth C. DePasqual Michigan Technological University Follow this and additional works at: https://digitalcommons.mtu.edu/etds Part of the Archaeological Anthropology Commons Copyright 2009 Seth C. DePasqual Recommended Citation DePasqual, Seth C., "Winning coal at 78° North : mining, contingency and the Chaîne Opératoire in old Longyear City", Master's Thesis, Michigan Technological University, 2009. https://doi.org/10.37099/mtu.dc.etds/308 Follow this and additional works at: https://digitalcommons.mtu.edu/etds Part of the Archaeological Anthropology Commons Winning Coal at 78˚ North: Mining, Contingency and the Chaîne Opératoire in Old Longyear City By Seth C. DePasqual A THESIS Submitted in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE IN INDUSTRIAL ARCHAEOLOGY MICHIGAN TECHNOLOGICAL UNIVERSITY 2009 This thesis, “Winning Coal at 78˚ North: Mining, Contingency and the Chaîne Opératoire in Old Longyear City” is hereby approved in partial fulfillment of the requirements for the Degree of MASTER OF SCIENCE IN INDUSTRIAL ARCHAEOLOGY. DEPARTMENT: Social Sciences Signatures: Thesis Advisor: ________________________________ Dr. Patrick E. Martin Department Chair: ______________________________ Dr. Patrick E. Martin Date: ______________________________ Acknowledgements This thesis would not have been possible without the encouraging words and guidance of my advisor Patrick Martin. His unremitting support saw me through a number of matters both academic and personal. I’d like to thank Carol MacLennan, who fostered my attention to socialized aspects of the Arctic Coal Company mining system and those related to the environment. -

Oppføringen Av Isfjord Radio, Automatiske Radiofyr Og Fyr Belysning Pa Svalbard 1946
Norges Svalbard- og Ishavs-undersøkelser Meddelelse nr. 67 Særtrykk av Norsk Geografisk Tidsskrift, bind XI, h. 5-6, 1947 REIDAR LYNGAAS OPPFØRINGEN AV ISFJORD RADIO, AUTOMATISKE RADIOFYR OG FYR BELYSNING PA SVALBARD 1946 A. W. B R Ø G G E R S B 0 K T RY K K E R I A/S - 0 S L 0 NORGES SVALBARD- OG ISHAVS-UNDERSØKELSER Observatoriegaten 1, Oslo MEDDELELSER Nr. I. PETTERSEN, K., ·Isforholdene i Nordishai•et i 1881 og 1882. Optrykk av avis artikler. Med en innledn. av A. Hoel. - Særtr. av Norsk Geogr. Tidsskr" · b. 1, h. 4. 1926. Kr. 1,00. [Utsolgt.] " 2. HOEL, A" Om ordninge11 av de territoriale krav på Svalbard. - Sertr. av Norsk Geogr. Tidsaler., b. 2, h. I. 1928. Kr. 1,60. [Utsolgt.] " 3. HOEL, fl." Suverenitetsspørsmålene i pofartraktene. - Særtr. av Nordmands Forbundet, årg. 21, h. 4 & 5. 1928. Kr. 1,00. [Utsolgt.] " 4. BROCH, 0. j., E. FJELD og A. HøYOAARD, På ski over den sydlige del av Spitsbergen. - Særtr. av Norsk Geogr. Tidsskr., b. 2, h. 3-4. 1928. Kr. 1,00. " 5 TANDBERG, ROLF S" Med hundespann på eftersøkning efter "ltalia"-folkene. - Særtr. av Norsk Geogr. Tidsskr. b. 2, h. 3-4. 1928. Kr. 2,20. " 6. KJÆR, IL Farvannsbeskrivelse over kysten av Bjørnøya. 1929. Kr. 1,60. " 7. NORGES SVALBARD- OG ISHAVS-UNDERSØKELSER, fan Mayen. En oversikt over øens natur, historie og bygning. - Særtr. av Norsk Geogr. Tidsskr., b. 2, h. 7. 1929. Kr. 1,60. !Utsolgt.] " 8. I. LID, JOHANNES, Mariskardet på. Svalbard. li. (SACHSEN, FRIDTJOV, Tidligere utforskning av områ.det mellem Isfjorden og Wijdebay på. -

Svalbard 2015–2016 Meld
Norwegian Ministry of Justice and Public Security Published by: Norwegian Ministry of Justice and Public Security Public institutions may order additional copies from: Norwegian Government Security and Service Organisation E-mail: [email protected] Internet: www.publikasjoner.dep.no KET T Meld. St. 32 (2015–2016) Report to the Storting (white paper) Telephone: + 47 222 40 000 ER RY M K Ø K J E L R I I Photo: Longyearbyen, Tommy Dahl Markussen M 0 Print: 07 PrintMedia AS 7 9 7 P 3 R 0 I 1 08/2017 – Impression 1000 N 4 TM 0 EDIA – 2 Svalbard 2015–2016 Meld. St. 32 (2015–2016) Report to the Storting (white paper) 1 Svalbard Meld. St. 32 (2015–2016) Report to the Storting (white paper) Svalbard Translation from Norwegian. For information only. Table of Contents 1 Summary ........................................ 5 6Longyearbyen .............................. 39 1.1 A predictable Svalbard policy ........ 5 6.1 Introduction .................................... 39 1.2 Contents of each chapter ............... 6 6.2 Areas for further development ..... 40 1.3 Full overview of measures ............. 8 6.2.1 Tourism: Longyearbyen and surrounding areas .......................... 41 2Background .................................. 11 6.2.2 Relocation of public-sector jobs .... 43 2.1 Introduction .................................... 11 6.2.3 Port development ........................... 44 2.2 Main policy objectives for Svalbard 11 6.2.4 Svalbard Science Centre ............... 45 2.3 Svalbard in general ........................ 12 6.2.5 Land development in Longyearbyen ................................ 46 3 Framework under international 6.2.6 Energy supply ................................ 46 law .................................................... 17 6.2.7 Water supply .................................. 47 3.1 Norwegian sovereignty .................. 17 6.3 Provision of services ..................... -

Sea Ice Cover in Isfjorden and Hornsund, Svalbard (2000-2014) from Remote Sensing Data S
Manuscript prepared for J. Name with version 2015/04/24 7.83 Copernicus papers of the LATEX class copernicus.cls. Date: 3 December 2015 Sea ice cover in Isfjorden and Hornsund, Svalbard (2000-2014) from remote sensing data S. Muckenhuber1, F. Nilsen2,3, A. Korosov1, and S. Sandven1 1Nansen Environmental and Remote Sensing Center (NERSC), Thormøhlensgate 47, 5006 Bergen, Norway 2University Centre in Svalbard (UNIS), P.O. Box 156, 9171 Longyearbyen, Norway 3Geophysical Institute, University of Bergen, P.O. Box 7800, 5020 Bergen, Norway Correspondence to: S. Muckenhuber ([email protected]) Abstract. A satellite database including 16 555 satellite images and ice charts displaying the area of Isfjorden, Hornsund and the Svalbard region has been established with focus on the time period 2000–2014. 3319 manual interpretations of sea ice conditions have been conducted, resulting in two time series dividing the area of Isfjorden and Hornsund into “Fast ice” (sea ice attached to the coast- 5 line), “Drift ice” and “Open water”. The maximum fast ice coverage of Isfjorden is > 40 % in the periods 2000–2005 and 2009–2011 and stays < 30 % in 2006–2008 and 2012–2014. Fast ice cover in Hornsund reaches > 40 % in all considered years, except for 2012 and 2014, where the maximum stays < 20 %. The mean seasonal cycles of fast ice in Isfjorden and Hornsund show monthly aver- aged values of less than 1 % between July and November and maxima in March (Isfjorden, 35.7 %) 10 and April (Hornsund, 42.1 %) respectively. A significant reduction of the monthly averaged fast ice coverage is found when comparing the time periods 2000–2005 and 2006–2014.