Coastal Erosion Reveals a Potentially Unique Oligocene and Possible Periglacial Sequence at Present-Day Sea Level in Port Davey, Remote South-West Tasmania
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

CLIMBING FEDERATION PEAK, SOUTH WEST TASMANIA Macquarie (University) Mountaineering Club Trip 1972 by Barbara Cameron Smith June 5, 2014
CLIMBING FEDERATION PEAK, SOUTH WEST TASMANIA Macquarie (University) Mountaineering Club trip 1972 By Barbara Cameron Smith June 5, 2014 Our anti clockwise route towards and up Federation Peak is depicted in orange above, with the exception of our detour off the loop to climb Burgess Bluff. We subsequently camped at Pineapple Flat, scrub bashed our way to Mount Picton, and eventually walked out to Blakes Opening along an unexpectedly civilized track. Map credit: Bill Filson 7 January 1972 We packed all our gear and then went shopping. We expect to be out for 7-10 days, and after packing the necessary food and excess, the food bill tallied 26 dollars for four, quite a lot of money. We went to local camping stores and got some extra equipment, then called in to chat with a few guys who could tell us something about the walk. We repacked everything after a counter lunch in a pub and off we went. We walked quite a way out of the main street of Hobart. Greg and I started hitching and were lucky, getting a lift with a guy who was going camping himself. I guess I was rather forward but I asked him if he’d mind picking up our two friends who were on the road already. He didn’t seem to mind, so we were all driven down to Geeveston. Had a few refreshments there and left details at the police station and gear at the council chambers. It was rather late to get a lift, it being 4.30 pm, but a local housewife drove all of us a few miles out of town. -

Background Report
Gordonvale World Heritage Area Reserve Background Report www.tasland.org.au Tasmanian Land Conservancy (2015). Gordonvale World Heritage Area Reserve Background Report. Tasmanian Land Conservancy, Tasmania Australia 7005. Copyright ©Tasmanian Land Conservancy The views expressed in this report are those of the Tasmanian Land Conservancy and not the Commonwealth Government, State Government or any other entity. This work is copyright. It may be reproduced for study, research or training purposes subject to an acknowledgment of the sources and no commercial usage or sale. Requests and enquires concerning reproduction and rights should be addressed to the Tasmanian Land Conservancy. Front Image: Gordonvale homestead (archive photo) and the Vale of Rasselas landscape © Grant Dixon Contact Address Tasmanian Land Conservancy PO Box 2112, Lower Sandy Bay, 827 Sandy Bay Road, Sandy Bay Tasmania, AUSTRALIA 7005 | p: 03 6225 1399 | www.tasland.org.au | Contents Page Acknowledgments 2 Acronyms and Abbreviations 3 INTRODUCTION 4 Gordonvale Fundraising Campaign 4 Location and Access 5 BIOREGIONAL AND LANDSCAPE CONTEXT 7 WORLD HERITAGE AREA STATUS 7 IUCN Protected Area Management Category 7 Legal Status and Compliance 8 NATURAL VALUES 9 GEO-CONSERVATION VALUES 12 WILDERNESS VALUES 12 INDIGENOUS VALUES 12 EUROPEAN HERITAGE 13 INVASIVE PESTS, WEEDS AND DISEASES 13 SCIENTIFIC STUDIES 13 MANAGEMENT STRATEGY 14 REFERENCES 15 APPENDIX A – Gordonvale ‘For Sale’ 16 APPENDIX B - Bushwalking tracks into Gordonvale 18 APPENDIX C - History of Ernie Bond at Gordonvale 21 1 Acknowledgements The Gordonvale World Heritage Area Reserve was secured by the Tasmanian Land Conservancy (TLC) in 2013 after a successful public fundraising campaign to purchase this private freehold block of land. -

3966 Tour Op 4Col
The Tasmanian Advantage natural and cultural features of Tasmania a resource manual aimed at developing knowledge and interpretive skills specific to Tasmania Contents 1 INTRODUCTION The aim of the manual Notesheets & how to use them Interpretation tips & useful references Minimal impact tourism 2 TASMANIA IN BRIEF Location Size Climate Population National parks Tasmania’s Wilderness World Heritage Area (WHA) Marine reserves Regional Forest Agreement (RFA) 4 INTERPRETATION AND TIPS Background What is interpretation? What is the aim of your operation? Principles of interpretation Planning to interpret Conducting your tour Research your content Manage the potential risks Evaluate your tour Commercial operators information 5 NATURAL ADVANTAGE Antarctic connection Geodiversity Marine environment Plant communities Threatened fauna species Mammals Birds Reptiles Freshwater fishes Invertebrates Fire Threats 6 HERITAGE Tasmanian Aboriginal heritage European history Convicts Whaling Pining Mining Coastal fishing Inland fishing History of the parks service History of forestry History of hydro electric power Gordon below Franklin dam controversy 6 WHAT AND WHERE: EAST & NORTHEAST National parks Reserved areas Great short walks Tasmanian trail Snippets of history What’s in a name? 7 WHAT AND WHERE: SOUTH & CENTRAL PLATEAU 8 WHAT AND WHERE: WEST & NORTHWEST 9 REFERENCES Useful references List of notesheets 10 NOTESHEETS: FAUNA Wildlife, Living with wildlife, Caring for nature, Threatened species, Threats 11 NOTESHEETS: PARKS & PLACES Parks & places, -

Characteristics of Interstate and Overseas Bushwalkers in the Arthur Ranges, South West Tasmania
CHARACTERISTICS OF INTERSTATE AND OVERSEAS BUSHWALKERS IN THE ARTHUR RANGES, SOUTH-WEST TASMANIA By Douglas A. Grubert & Lorne K. Kriwoken RESEARCH REPORT RESEARCH REPORT SERIES The primary aim of CRC Tourism's research report series is technology transfer. The reports are targeted toward both industry and government users and tourism researchers. The content of this technical report series primarily focuses on applications, but may also advance research methodology and tourism theory. The report series titles relate to CRC Tourism's research program areas. All research reports are peer reviewed by at least two external reviewers. For further information on the report series, access the CRC website [www.crctourism.com.au]. EDITORS Prof Chris Cooper University of Queensland Editor-in-Chief Prof Terry De Lacy CRC for Sustainable Tourism Chief Executive Prof Leo Jago CRC for Sustainable Tourism Director of Research National Library of Australia Cataloguing-in-Publication Data Grubert, Douglas. Characteristics of interstate and overseas bushwalkers in the Arthur Ranges, South West Tasmania. Bibliography. ISBN 1 876685 83 2. 1. Hiking - Research - Tasmania - Arthur Range. 2. Hiking - Tasmania - Arthur Range - Statistics. 3. National parks and reserves - Public use - Tasmania - Arthur Range. I. Kriwoken, Lorne K. (Lorne Keith). II. Cooperative Research Centre for Sustainable Tourism. III. Title. 796.52209946 © 2002 Copyright CRC for Sustainable Tourism Pty Ltd All rights reserved. No parts of this report may be reproduced, stored in a retrieval system or transmitted in any form or by means of electronic, mechanical, photocopying, recording or otherwise without the prior permission of the publisher. Any enquiries should be directed to Brad Cox, Director of Communications or Trish O’Connor, Publications Manager to [email protected]. -

A Review of Geoconservation Values
Geoconservation Values of the TWWHA and Adjacent Areas 3.0 GEOCONSERVATION AND GEOHERITAGE VALUES OF THE TWWHA AND ADJACENT AREAS 3.1 Introduction This section provides an assessment of the geoconservation (geoheritage) values of the TWWHA, with particular emphasis on the identification of geoconservation values of World Heritage significance. This assessment is based on: • a review (Section 2.3.2) of the geoconservation values cited in the 1989 TWWHA nomination (DASETT 1989); • a review of relevant new scientific data that has become available since 1989 (Section 2.4); and: • the use of contemporary procedures for rigorous justification of geoconservation significance (see Section 2.2) in terms of the updated World Heritage Criteria (UNESCO 1999; see this report Section 2.3.3). In general, this review indicates that the major geoconservation World Heritage values of the TWWHA identified in 1989 are robust and remain valid. However, only a handful of individual sites or features in the TWWHA are considered to have World Heritage value in their own right, as physical features considered in isolation (eg, Exit Cave). In general it is the diversity, extent and inter-relationships between numerous features, sites, areas or processes that gives World Heritage significance to certain geoheritage “themes” in the TWWHA (eg, the "Ongoing Natural Geomorphic and Soil Process Systems" and “Late Cainozoic "Ice Ages" and Climate Change Record” themes). This "wholistic" principle under-pinned the 1989 TWWHA nomination (DASETT 1989, p. 27; see this report Section 2.3.2), and is strongly supported by the present review (see discussion and justification of this principle in Section 2.2). -

CHANGES in SOUTHWESTERN TASMANIAN FIRE REGIMES SINCE the EARLY 1800S
Papers and Proceedings o/the Royal Society o/Tasmania, Volume 132, 1998 IS CHANGES IN SOUTHWESTERN TASMANIAN FIRE REGIMES SINCE THE EARLY 1800s by Jon B. Marsden-Smedley (with five tables and one text-figure) MARSDEN-SMEDLEY, ].B., 1998 (31:xii): Changes in southwestern Tasmanian fire regimes since the early 1800s. Pap.Proc. R. Soc. Tasm. 132: 15-29. ISSN 0040-4703. School of Geography and Environmental Studies, University of Tasmania, GPO Box 252-78, Hobart, Tasmania, Australia 7001. There have been major changes in the fire regime of southwestern Tasmania over the past 170 years. The fire regime has changed from an Aboriginal fire regime of frequent low-intensity fires in buttongrass moorland (mostly in spring and autumn) with only the occasional high-intensity forest fire, to the early European fire regime of frequent high-intensity fires in all vegetation types, to a regime of low to medium intensity buttongrass moorland fires and finally to the current regime of few fires. These changes in the fire regime resulted in major impacts to the region's fire-sensitive vegetation types during the early European period, while the current low fire frequency across much of southwestern Tasmania has resulted in a large proportion of the region's fire-adapted buttongrass moorland being classified as old-growth. These extensive areas of old-growth buttongrass moorland mean that the potential for another large-scale ecologically damaging wildfire is high and, to avoid this, it would be better to re-introduce a regime oflow-intensity fires into the region. Key Words: fire regimes, fire management, southwestern Tasmania, Aboriginal fire, history. -

Conservation of Natural Wilderness Values in the Port Davey Marine and Estuarine Protected Area, South-Western Tasmania
AQUATIC CONSERVATION: MARINE AND FRESHWATER ECOSYSTEMS Aquatic Conserv: Mar. Freshw. Ecosyst. 20: 297–311 (2010) Published online 3 December 2009 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/aqc.1079 Conservation of natural wilderness values in the Port Davey marine and estuarine protected area, south-western Tasmania GRAHAM J. EDGARa,b,Ã, PETER R. LASTc, NEVILLE S. BARRETTb, KAREN GOWLETT-HOLMESc, MICHAEL DRIESSENd and PETER MOONEYe aAquenal Pty Ltd, GPO Box 828, Hobart, Tasmania, Australia 7001 bTasmanian Aquaculture and Fisheries Institute, University of Tasmania, GPO Box 252-49, Hobart, Tasmania, Australia 7001 cCSIRO Divison of Marine and Atmospheric Research, Castray Esplanade, Hobart, Tasmania, Australia 7000 dDepartment of Primary Industries and Water, Hobart, Tasmania, Australia 7000 eParks and Wildlife Service, Hobart, Tasmania, Australia 7000 ABSTRACT 1. Port Davey and associated Bathurst Harbour in south-western Tasmania represent one of the world’s most anomalous estuarine systems owing to an unusual combination of environmental factors. These include: (i) large uninhabited catchment protected as a National Park; (ii) ria geomorphology but with fjord characteristics that include a shallow entrance and deep 12-km long channel connecting an almost land-locked harbour to the sea; (iii) high rainfall and riverine input that generate strongly-stratified estuarine conditions, with a low-salinity surface layer and marine bottom water; (iv) a deeply tannin-stained surface layer that blocks light penetration to -

Nowhere Else on Earth
Nowhere Else on Earth: Tasmania’s Marine Natural Values Environment Tasmania is a not-for-profit conservation council dedicated to the protection, conservation and rehabilitation of Tasmania’s natural environment. Australia’s youngest conservation council, Environment Tasmania was established in 2006 and is a peak body representing over 20 Tasmanian environment groups. Prepared for Environment Tasmania by Dr Karen Parsons of Aquenal Pty Ltd. Report citation: Parsons, K. E. (2011) Nowhere Else on Earth: Tasmania’s Marine Natural Values. Report for Environment Tasmania. Aquenal, Tasmania. ISBN: 978-0-646-56647-4 Graphic Design: onetonnegraphic www.onetonnegraphic.com.au Online: Visit the Environment Tasmania website at: www.et.org.au or Ocean Planet online at www.oceanplanet.org.au Partners: With thanks to the The Wilderness Society Inc for their financial support through the WildCountry Small Grants Program, and to NRM North and NRM South. Front Cover: Gorgonian fan with diver (Photograph: © Geoff Rollins). 2 Waterfall Bay cave (Photograph: © Jon Bryan). Acknowledgements The following people are thanked for their assistance The majority of the photographs in the report were with the compilation of this report: Neville Barrett of the generously provided by Graham Edgar, while the following Institute for Marine and Antarctic Studies (IMAS) at the additional contributors are also acknowledged: Neville University of Tasmania for providing information on key Barrett, Jane Elek, Sue Wragge, Chris Black, Jon Bryan, features of Tasmania’s marine -

Reimagining the Visitor Experience of Tasmania's Wilderness World
Reimagining the Visitor Experience of Tasmania’s Wilderness World Heritage Area Ecotourism Investment Profile Reimagining the Visitor Experience of Tasmania’s Wilderness World Heritage Area: Ecotourism Investment Profile This report was commissioned by Tourism Industry Council Tasmania and the Cradle Coast Authority, in partnership with the Tasmanian Government through Tourism Tasmania and the Tasmanian Parks and Wildlife Service. This report is co-funded by the Australian Government under the Tourism Industry Regional Development Fund Grants Programme. This report has been prepared by EC3 Global, TRC Tourism and Tourism Industry Council Tasmania. Date prepared: June 2014 Design by Halibut Creative Collective. Disclaimer The information and recommendations provided in this report are made on the basis of information available at the time of preparation. While all care has been taken to check and validate material presented in this report, independent research should be undertaken before any action or decision is taken on the basis of material contained in this report. This report does not seek to provide any assurance of project viability and EC3 Global, TRC Tourism and Tourism Industry Council Tasmania accept no liability for decisions made or the information provided in this report. Cover photo: Huon Pine Walk Corinna The Tarkine - Rob Burnett & Tourism Tasmania Contents Background...............................................................2 Reimagining the Visitor Experience of the TWWHA .................................................................5 -

The Lake Pedder Decision
Hrasky, S. , & Jones, M. J. (2016). Lake Pedder: Accounting, environmental decision-making, nature and impression management. Accounting Forum, 40(4), 285-299. https://doi.org/10.1016/j.accfor.2016.06.005 Peer reviewed version License (if available): CC BY-NC-ND Link to published version (if available): 10.1016/j.accfor.2016.06.005 Link to publication record in Explore Bristol Research PDF-document This is the accepted author manuscript (AAM). The final published version (version of record) is available online via Elsevier at http://dx.doi.org/10.1016/j.accfor.2016.06.005. Please refer to any applicable terms of use of the publisher. University of Bristol - Explore Bristol Research General rights This document is made available in accordance with publisher policies. Please cite only the published version using the reference above. Full terms of use are available: http://www.bristol.ac.uk/red/research-policy/pure/user-guides/ebr-terms/ Lake Pedder: Accounting, Environmental Decision-Making, nature and impression management Sue Hrasky and Michael Jones University of Tasmania, University of Bristol Acknowledgements We wish to thank participants at the 15th Financial Reporting and Business Communication Conference, Bristol, July 2011, the 23rd International Congress on Social and Environmental Accounting Research (CSEAR), St Andrews, September 2011 and the 10th CSEAR Australasian Conference, Launceston, December 2011, for their helpful comments and suggestions. My thanks also to Claire Horner for her help collecting data. Finally, I would like to thank Glen Lehman and an anonymous reviewer Corresponding Author Department of Accounting and Finance University of Bristol 8 Woodland Road, Bristol BS8 1TN, UK Email: [email protected] Phone: +44 (0)117 33 18286 Lake Pedder: Accounting, Environmental Decision-Making, Nature and Impression Management Abstract This paper looks at the role of accounting in a major environmental infrastructural project the flooding of Lake Pedder in Tasmania in the 1960s. -

Melaleuca Site and Rehabilitation Plan 2014
Melaleuca Site and Rehabilitation Plan Southwest National Park Tasmanian Wilderness World Heritage Area Melaleuca Site and Rehabilitation Plan This site and rehabilitation plan applies to Melaleuca which lies in the Southwest National Park and the Tasmanian Wilderness World Heritage Area. It applies to the Melaleuca Visitor Services Zone as well as the former Rallinga mining lease area and nearby sites used for recreational, conservation or operational purposes. The plan has been prepared, as agreed with the Commonwealth Government, to facilitate mining rehabilitation works; to improve visitors’ experiences in the Melaleuca area; and, to guide site management. Peter Mooney General Manager Tasmania Parks and Wildlife Service ISBN 978-0-9923628-1-2 (print version) ISBN 978-0-9923628-2-9 (pdf version) © State of Tasmania 2014 Cover photo: Moth Creek by A.Turbett Published by: Parks and Wildlife Service Department of Primary Industries, Parks, Water and Environment GPO Box 1751 Hobart TASMANIA 7001 Melaleuca Site and Rehabilitation Plan Contents Visitor Experience Statement ................................................................................................ 1 Summary ................................................................................................................................... 3 1. Introduction and reserve management framework ...................................................... 4 1.1 Significance of Melaleuca ................................................................................................................................. -
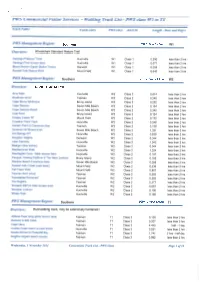
Walking Track List - PWS Class Wl to T4
PWS Commercial Visitor Services - Walking Track List - PWS class Wl to T4 Track Name FieldCentre PWS class AS2156 Length - Kms and Days PWS Management Region: Southern PWS Track Class: VV1 Overview: Wheelchair Standard Nature Trail Hastings Platypus Track Huonville W1 Class 1 0.290 less than 2 hrs Hastings Pool access track Huonville W1 Class 1 0.077 less than 2 hrs Mount Nelson Signal Station Tracks Derwent W1 Class 1 0.059 less than 2 hrs Russell Falls Nature Walk Mount Field W1 Class 1 0.649 less than 2 hrs PWS Management Region: Southern PWS Track Class: W2 Overview: Standard Nature Trail Arve Falls Huonville W2 Class 2 0.614 less than 2 hrs Blowhole circuit Tasman W2 Class 2 0.248 less than 2 hrs Cape Bruny lighthouse Bruny Island W2 Class 2 0.252 less than 2 hrs Cape Deslacs Seven Mile Beach W2 Class 2 0.154 less than 2 hrs Cape Deslacs Beach Seven Mile Beach W2 Class 2 0.345 less than 2 hrs Coal Point Bruny Island W2 Class 2 0.124 less than 2 hrs Creepy Crawly NT Mount Field W2 Class 2 0.175 less than 2 hrs Crowther Point Track Huonville W2 Class 2 0.248 less than 2 hrs Garden Point to Carnarvon Bay Tasman W2 Class 2 3.138 less than 2 hrs Gordons Hill fitness track Seven Mile Beach W2 Class 2 1.331 less than 2 hrs Hot Springs NT Huonville W2 Class 2 0.839 less than 2 hrs Kingston Heights Derwent W2 Class 2 0.344 less than 2 hrs Lake Osbome Huonville W2 Class 2 1.042 less than 2 hrs Maingon Bay lookout Tasman W2 Class 2 0.044 less than 2 hrs Needwonnee Walk Huonville W2 Class 2 1.324 less than 2 hrs Newdegate Cave - Main access