Control and Efficiency of Digestive Function of Marine Fish Larvae
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Coffee and Its Effect on Digestion
Expert report Coffee and its effect on digestion By Dr. Carlo La Vecchia, Professor of Medical Statistics and Epidemiology, Dept. of Clinical Sciences and Community Health, Università degli Studi di Milano, Italy. Contents 1 Overview 2 2 Coffee, a diet staple for millions 3 3 What effect can coffee have on the stomach? 4 4 Can coffee trigger heartburn or GORD? 5 5 Is coffee associated with the development of gastric or duodenal ulcers? 6 6 Can coffee help gallbladder or pancreatic function? 7 7 Does coffee consumption have an impact on the lower digestive tract? 8 8 Coffee and gut microbiota — an emerging area of research 9 9 About ISIC 10 10 References 11 www.coffeeandhealth.org May 2020 1 Expert report Coffee and its effect on digestion Overview There have been a number of studies published on coffee and its effect on different areas of digestion; some reporting favourable effects, while other studies report fewer positive effects. This report provides an overview of this body of research, highlighting a number of interesting findings that have emerged to date. Digestion is the breakdown of food and drink, which occurs through the synchronised function of several organs. It is coordinated by the nervous system and a number of different hormones, and can be impacted by a number of external factors. Coffee has been suggested as a trigger for some common digestive complaints from stomach ache and heartburn, through to bowel problems. Research suggests that coffee consumption can stimulate gastric, bile and pancreatic secretions, all of which play important roles in the overall process of digestion1–6. -

Study Guide Medical Terminology by Thea Liza Batan About the Author
Study Guide Medical Terminology By Thea Liza Batan About the Author Thea Liza Batan earned a Master of Science in Nursing Administration in 2007 from Xavier University in Cincinnati, Ohio. She has worked as a staff nurse, nurse instructor, and level department head. She currently works as a simulation coordinator and a free- lance writer specializing in nursing and healthcare. All terms mentioned in this text that are known to be trademarks or service marks have been appropriately capitalized. Use of a term in this text shouldn’t be regarded as affecting the validity of any trademark or service mark. Copyright © 2017 by Penn Foster, Inc. All rights reserved. No part of the material protected by this copyright may be reproduced or utilized in any form or by any means, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without permission in writing from the copyright owner. Requests for permission to make copies of any part of the work should be mailed to Copyright Permissions, Penn Foster, 925 Oak Street, Scranton, Pennsylvania 18515. Printed in the United States of America CONTENTS INSTRUCTIONS 1 READING ASSIGNMENTS 3 LESSON 1: THE FUNDAMENTALS OF MEDICAL TERMINOLOGY 5 LESSON 2: DIAGNOSIS, INTERVENTION, AND HUMAN BODY TERMS 28 LESSON 3: MUSCULOSKELETAL, CIRCULATORY, AND RESPIRATORY SYSTEM TERMS 44 LESSON 4: DIGESTIVE, URINARY, AND REPRODUCTIVE SYSTEM TERMS 69 LESSON 5: INTEGUMENTARY, NERVOUS, AND ENDOCRINE S YSTEM TERMS 96 SELF-CHECK ANSWERS 134 © PENN FOSTER, INC. 2017 MEDICAL TERMINOLOGY PAGE III Contents INSTRUCTIONS INTRODUCTION Welcome to your course on medical terminology. You’re taking this course because you’re most likely interested in pursuing a health and science career, which entails proficiencyincommunicatingwithhealthcareprofessionalssuchasphysicians,nurses, or dentists. -
The Role of Lactic Acid in Gastric Digestion
[Reprinted from The Medical News, December 30, 1893.] THE ROLE OF LACTIC ACID IN GASTRIC DIGESTIOA ALLEN A. JONES, M.D., CLINICAL INSTRUCTOR IN MEDICINE AND INSTRUCTOR IN PRACTICE, MEDICAL DEPARTMENT, UNIVERSITY OF BUFFALO. Lactic acid is present in the stomach under nor- mal conditions from thirty to forty minutes after a test-meal composed of a roll and water or of chopped lean beef, dry bread, and water. At the expiration of that time lactic acid should entirely disappear from the stomach-contentsand free hydro- chloric acid alone should prevail. During the first thirty or forty minutes after meals the digestion of starches and albuminoids progresses quite rapidly, as may be proved by finding the middle-products and end-products of gastric digestion present, so that the presence of free lactic acid does not pro- hibit digestion. As soon as food enters the healthy stomach the secretion of hydrochloric acid is excited and it increases in amount until the production of lactic acid is checked. The exact origin of lactic acid in the healthy stomach is still a matter of debate. It may arise wholly from fermentation, or from the combination of some food-product with a secretion 1 Read before the Buffalo Academy of Medicine, November x 4) 1893. 2 from the gastric glandules, or from the gastric mucosa as a distinct secretion, although electric stimulation of the gastric glandules excites the se- cretion of hydrochloric acid and not of lactic acid. I think it arises largely from fermentation of the food, as its amount is usually proportionate to the amount of starchy, saccharine, and milk foods taken. -

Intestinal and Liver Morphometry of the Yellow Tail Tetra (Astyanax Altiparanae) Fed with Oregano Oil
Anais da Academia Brasileira de Ciências (2016) 88(2): 911-922 (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 http://dx.doi.org/10.1590/0001-3765201620150202 www.scielo.br/aabc Intestinal and liver morphometry of the Yellow Tail Tetra (Astyanax altiparanae) fed with oregano oil POLLYANNA M.F. FERREIRA, DÉBORA W. CALDAS, ANA LÚCIA SALARO, SIRLENE S.R. SARTORI, JERUSA M. OLIVEIRA, ALEX J.S. CARDOSO and JENER A.S. ZUANON Departamento de Biologia Animal, Universidade Federal de Viçosa/UFV, Av. PH Rolfs, s/n, 36570-900 Viçosa, MG, Brasil Manuscript received on April 6, 2015; accepted for publication on August 20, 2015 ABSTRACT This study aimed to evaluate the effect of oregano oil on the intestinal and liver morphometry of yellow tail tetra, Astyanax altiparanae. Fish (1.46 ± 0.09 g) were kept in a 60-L aquaria, at a stocking density of 0.5 fi sh L-1. Six diets containing varying amounts of oregano oil were evaluated (0.0; 0.5; 1.0; 1.5; 2.0 and 2.5 g of oregano oil kg-1). At the end of 90 days, the fi sh were euthanised. Four intestines and four livers were collected per treatment, which were fi xed in Bouin and embedded in resin. For height and width folds, the absorption surface area and thickness of the muscular layer a positive linear effect of oregano oil was observed. A decrescent linear effect on the total number of goblet cells was also observed. For the cytoplasmic percentage of hepatocytes and liver glycogen, a positive linear effect of oregano oil was observed. -

New Developments in Goblet Cell Mucus Secretion and Function
REVIEW nature publishing group New developments in goblet cell mucus secretion and function GMH Birchenough1, MEV Johansson1, JK Gustafsson1, JH Bergstro¨m1 and GC Hansson1 Goblet cells and their main secretory product, mucus, have long been poorly appreciated; however, recent discoveries have changed this and placed these cells at the center stage of our understanding of mucosal biology and the immunology of the intestinal tract. The mucus system differs substantially between the small and large intestine, although it is built around MUC2 mucin polymers in both cases. Furthermore, that goblet cells and the regulation of their secretion also differ between these two parts of the intestine is of fundamental importance for a better understanding of mucosal immunology. There are several types of goblet cell that can be delineated based on their location and function. The surface colonic goblet cells secrete continuously to maintain the inner mucus layer, whereas goblet cells of the colonic and small intestinal crypts secrete upon stimulation, for example, after endocytosis or in response to acetyl choline. However, despite much progress in recent years, our understanding of goblet cell function and regulation is still in its infancy. THE INTESTINE system of mucus covering the epithelium. There is a The gastrointestinal tract is a remarkable organ. Not only can it two-layered mucus system in the stomach and colon and a digest most of our food into small components, but it is also single-layered mucus in the small intestine.5 The mucus layers filled with kilograms of microbes that live in stable equilibrium in these three regions perform their protective function using with us and our immune system. -

Cf Facts — the Digestive System
Beginning CF Care — CF FACTS — THE DIGESTIVE SYSTEM CF FACTS — THE DIGESTIVE SYSTEM THE GI TRACT THE PANCREAS AND LIVER the small intestine through a series Digestion * takes place in the Two other organs found in the of tubes. When there is food in gastrointestinal (GI) tract .* The abdomen * (belly) help with the small intestine, the enzymes GI tract is also called the digestive digestion: the pancreas * and the help break the food down so it tract. The GI tract is basically a liver .* The pancreas is an organ can be absorbed and used by the long tube that begins with the that sits in the upper abdomen body. The pancreas also produces mouth and continues through the behind the stomach. The pancreas insulin * that helps the body use esophagus ,* stomach, small, and produces enzymes * or special glucose ,* a sugar that comes from large intestines .* (The small and proteins that break down fat * and the digestion of carbohydrates .* large intestines together are about protein * in food. These enzymes Insulin is released into blood that 25 feet long!) The GI tract ends at include lipase ,* protease ,* and passes through the pancreas. the rectum * and anus .* amylase .* The enzymes pass into The liver is an organ that sits in the upper right side of the abdomen. The gallbladder * is attached to the liver and helps store extra bile * fluid that is made by the liver. The liver and gallbladder are connected to the small intestine by a tube. The liver does many things for the body. Bile fluid is sent from the liver to the small intestine to help with digestion. -

GLOSSARYGLOSSARY Medical Terms Common to Hepatology
GLOSSARYGLOSSARY Medical Terms Common to Hepatology Abdomen (AB-doh-men): The area between the chest and the hips. Contains the stomach, small intestine, large intestine, liver, gallbladder, pancreas and spleen. Absorption (ub-SORP-shun): The way nutrients from food move from the small intestine into the cells in the body. Acetaminophen (uh-seat-uh-MIN-oh-fin): An active ingredient in some over-the-counter fever reducers and pain relievers, including Tylenol. Acute (uh-CUTE): A disorder that has a sudden onset. Alagille Syndrome (al-uh-GEEL sin-drohm): A condition when the liver has less than the normal number of bile ducts. It is associated with other characteristics such as particular facies, abnormal pulmonary artery and abnormal vertebral bodies. Alanine Aminotransferase or ALT (AL-ah-neen uh-meen-oh-TRANZ-fur-ayz): An enzyme produced by hepatocytes, the major cell types in the liver. As cells are damaged, ALT leaks out into the bloodstream. ALT levels above normal may indicate liver damage. Albumin (al-BYEW-min): A protein that is synthesized by the liver and secreted into the blood. Low levels of albumin in the blood may indicate poor liver function. Alimentary Canal (al-uh-MEN-tree kuh-NAL): See Gastrointestinal (GI) Tract. Alkaline Phosphatase (AL-kuh-leen FOSS-fuh-tayz): Proteins or enzymes produced by the liver when bile ducts are blocked. Allergy (AL-ur-jee): A condition in which the body is not able to tolerate or has a reaction to certain foods, animals, plants, or other substances. Amino Acids (uh-MEE-noh ASS-udz): The basic building blocks of proteins. -
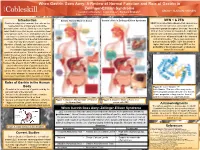
A Review of Normal Function and Role of Gastrin in Zollinger-Ellison
When Gastrin Goes Awry: A Review of Normal Function and Role of Gastrin in Zollinger-Ellison Syndrome Leigha LaTourette1, Janel Cross2, Barbara Brabetz2 Department of Plant and Animal Science1, Department of Natural Sciences and Mathematics2 Introduction Gastrin: Normal Mode of Action Gastrin’s Role in Zollinger-Ellison Syndrome MEN 1 & ZES Gastrin is a digestive hormone that acts on the MEN1 is an inheritable disease that causes over neuroendocrine and parietal cells of the secretion of hormones via tumors on the gastrointestinal tract to ultimately secrete gastric endocrine glands throughout the body.4 Around a acid. Gastrin secretion begins even before food 30% of these tumors are found to be malignant. is consumed, as the mere anticipation of a meal ZES is commonly associated with the MEN1 due can lead to a series of events ending in the to the presence of tumors found in the stomach creation of the gastric acid needed to breakdown and small intestine.4 These gastrointestinal foods. Not only does Gastrin support the tumors caused by MEN 1 give the person a digestion of foods, but it also stimulates growth, higher likelihood of contracting ZES, as the secretion, blood flow, and acts as a defense probability of having pancreatic or duodenal mechanism against bacteria in the tumors is increased.4 gastrointestinal system. When the production of Gastrin becomes overt, major consequences like that of Zollinger Ellison Syndrome (ZES). ZES is an extremely rare disease occurring in people between the ages of 30-60.4 ZES is related to the uncontrolled secretion of gastrin due to the presence of certain pancreatic or duodenal tumors. -

Pancreas and Fat/Lipid Digestion
Exocrine Pancreas Physiology Pancreatic Anatomy Pancreatic secretion Pancreatic enzymes Daniel S. Kamin MD Boston Children’s Hospital [email protected] Content Reviewers: Sohail Z. Husain, MD Veronique Morinville MD, FRCP(C) NASPGHAN Physiology Education Series Series Editors: Christine Waasdorp Hurtado, MD, MSCS, FAAP [email protected] Daniel Kamin, MD [email protected] Learning Objectives • Understand the normal development and anatomy of the pancreas • Understand the stimuli and cellular factors giving rise to pancreatic secretion • Know the mechanisms by which pancreatic enzymes are activated and remain functional • Be aware of age-related deficiency in exocrine pancreatic function Night blindness • A young man with morbid obesity undergoes roux-en-y gastric bypass. • 1 year later he notices that in the evening he falls down his stairs. • Vitamin A deficiency is diagnosed. Understanding the physiology of pancreatic and bile secretion, Used with permission intraluminal lipolysis, and micellar function explains why this happens! http://www.citelighter.com/science/surgery/knowledgecards/gastric-bypass Pancreas Physiology Overview • Bulk of bicarbonate secretion (more than what secreted in bile and from duodenum) • Enzymes for intra- luminal digestion • Secretin and CCK regulate • Maturational pancreatic insufficiency Used with permission Image from http://www.aboutcancer.com/pancreas1.htm Pancreatic Development • See Embryology and Anatomy of the Gastrointestinal Tract Pancreatic Microanatomy Pancreatic Acinar Secretory Products A • Proteases • Trypsinogen* • Chymotrypsinogen* A • Proelastase* • Procarboxypeptidase* • Procarboxypeptidase B* • Amylolytic enzyme • Amylase A • Lipases • Lipase • B Nonspecific esterase • Prophospholipase A2* • Nucleases • Deoxyribonuclease • Ribonuclease • Others • Pro-colipase* • Trypsin inhibitors A. Exocrine pancreas-- ascinar cells filled • Monitor peptid with secretory granules, cuboidal duct Stored and secreted in inactive form cells secrete bicarbonate-rich fluid B. -

A History of Gastric Secretion and Digestion a History of Gastric Secretion and Digestion Experimental Studies to 1975
A History of Gastric Secretion and Digestion A History of Gastric Secretion and Digestion Experimental Studies to 1975 HORACE W. DAVENPORT William Beaumont Professor of Physiology Emeritus The University of Michigan Springer New Y ork 1992 Copyright © 1992 by the American Physiological Society Originally published by American Physiological Society in 1992 Softcoverreprint of the bardeover 1st edition 1992 All rights reserved. No partoftbis publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, electronic, mechanical, photocopying, recording, or otherwise, without the prior permission ofOxford University Press. Library ofCongress Cataloging-in-Publication Data Davenport, Horace Willard, 1912- A history of gastric secretion and digestion : experimental studiesto 1975 I Horace W. Davenport. p. cm. lncludes bibliographical references and index. ISBN 978-1-4614-7602-3 (eBook) DOI 10.1007/978-1-4614-7602-3 I. Gastroenterology-History. 2. Gastric-Secretion-Research-History. 3. Digestion-Research-History. I. Title. [DNLM: I. Digestion. 2. Gastric Acid-secretion. 3. Gastroenterology-history. 4. Research-history. 5. Stomach-chemistry. 6. Stomach-physiology. Wlll.l D247h] QP145.D325 1992 612.3'2'072-dc20 DNLM/DLC for Library ofCongress 91-31832 987654321 For Charles F. Code, known to every gastroenterologist as "Charlie Code" and as their preeminent physiologist for the last fifty years Preface For centuries men speculated about the process of gastric digestion, but Iate in the eighteenth and early in the nineteenth centuries physiologists, both physicians and laymen, began to accumulate experimental evidence about its nature. At the same time, others discovered that the stomach is capable of secreting a strong mineral acid, and the questions of how that secretion is produced and how it is controlled became enduring problems. -

The Digestive System
THE DIGESTIVE SYSTEM COMPILED BY HOWIE BAUM DIGESTIVE SYSTEM People are probably more aware of their digestive system than of any other system, not least because of its frequent messages. Hunger, thirst, appetite, gas ☺, and the frequency and nature of bowel movements, are all issues affecting daily life. The Digestive Tract • Six Functions of the Digestive System 1. Ingestion 2. Mechanical processing 3. Digestion 4. Secretion 5. Absorption 6. Excretion The Digestive Tract • Ingestion – Occurs when materials enter digestive tract via the mouth • Mechanical Processing – Crushing and shearing – Makes materials easier to propel along digestive tract • Digestion – The chemical breakdown of food into small organic fragments for absorption by digestive epithelium The Digestive Tract • Secretion – Is the release of water, acids, enzymes, buffers, and salts – By epithelium of digestive tract – By glandular organs • Absorption – Movement of organic substrates, electrolytes, vitamins, and water – Across digestive epithelium tissue – Into the interstitial fluid of digestive tract • Excretion – Removal of waste products from body fluids – Process called defecation removes feces AN INTRODUCTION TO THE DIGESTIVE SYSTEM • The Digestive Tract • Also called the gastrointestinal (GI) tract or alimentary canal • Is a muscular tube • Extends from our mouth to the anus • Passes through the pharynx, esophagus, stomach, and small and large intestines The digestive system is one of the most clearly defined in the body. It consists of a long passageway, the digestive -

Gastrointestinal Glossary of Terms
Gastrointestinal Glossary of Terms Abdominoperineal resection Surgical removal of the anus, rectum and sigmoid colon, resulting in the need for a permanent colostomy. Adenoma Glandular lesion thought to be the precursor to colorectal cancer. Adhesion A band of scar tissue that connects two surfaces of the body that are normally separate. Air contrast barium enema An X-ray examination of the entire large intestine (colon) and rectum in which barium and air are introduced gradually into the colon by a rectal tube. This test is recommended along with flexible sigmoidoscopy every five years, starting at age 50, to screen for colorectal cancer and polyps. Anal fissure A split or crack in the lining of the anal opening, usually caused by the passage of very hard or watery stools. Anastomosis A surgical joining of two ducts, blood vessels or bowel segments to allow flow from one to the other. Aneurysm The abnormal enlargement or bulging of a blood vessel, caused by damage or weakness in the blood vessel wall. Angiogram A technique that uses dye to highlight blood vessels. Anus The opening at one end of the digestive tract from which waste is expelled. Appendectomy Surgical removal of the appendix to treat appendicitis. Appendicitis Inflammation of the appendix that requires immediate medical attention. Appendix A small, finger-like tube located where the large and small intestine join. It has no known function. Ascites Fluid in the abdomen. Banding A technique via endoscopy that the dilated blood vessels in the esophagus can be removed by putting rubber bands on and eventually fall off to the disappearance of those vessels.