Parasite Infection
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Grand Teton National Park
GRAND TETON NATIONAL PARK • WTO MING * UNITED STATES DEPARTMENT OF THE INTERIOR NATIO NAL PARK SERVICE Grand Teton [WYOMING] National Park United States Department of the Interior Harold L. Ickes, Secretary NATIONAL PARK SERVICE Arno B. Cammerer, Director UNITED STATES GOVERNMENT PRINTING OFFICE WASHINGTON : 1936 Rules and Regulations -I-HE PARK regulations are designed for the protection of the natural Contents beauties as well as for the comfort and convenience of visitors. The follow ing synopsis is for the general guidance of visitors, who are requested to assist in the administration of the park by observing them. Copies of the complete rules and regulations promulgated by the Secretary of the Interior Page for the government of the park may be obtained at the office of the super History of the Region 3 intendent and at other points of concentration throughout the park. Geographic Features 7 The destruction, injury, defacement, or disturbance of any buildings, Teton Range 7 signs, equipment, trees, flowers, vegetation, rocks, minerals, animal, bird, Jackson Hole 9 or other life is prohibited. The Work of Glaciers 9 Camps must be kept clean. Rubbish and garbage should be burned. Trails 13 Refuse should be placed in cans provided for this purpose. If no cans are Mountain Climbing 14 provided where camp is made, refuse should be buried. Wildlife 18 Do not throw paper, lunch refuse, or other trash on the roads and trails. Trees and Plants 21 Carry until the same can be burned in camp or placed in receptacle. Naturalist Service 23 Fires shall be lighted only when necessary and when no longer needed Fishing 24 shall be completely extinguished. -

A Brief History of the Trails of Grand Teton National Park 55
Pritchard: A Brief History of the Trails of Grand Teton National Park 55 A BRIEF HISTORY OF THE TRAILS OF GRAND TETON NATIONAL PARK JAMES A. PRITCHARD IOWA STATE UNIVERSITY AMES ABSTRACT reconstructed during the MISSION 66 era, but some of the stone stairs along the way from the boat dock This project investigated the history of the to Hidden Falls date back to the CCC era. backcountry trail system in Grand Teton National Park (GTNP). In cooperation with GTNP Cultural Walking on a beautiful mountain path, one Resources and the Western Center for Historic might never guess the extensive preparation of rock Preservation in GTNP, we located records describing materials (expediting drainage) that is required before the early development of the trail system. Only a few the surface ―treadway‖ is laid down (Barter et al. historical records describe or map the exact location 2006). In fact, trails are significant engineering of early trails, which prove useful when relocating achievements that need constant care and upkeep, trails today. The paper trail becomes quite rich, including annual clearance of vegetation and the however, in revealing the story behind the practical occasional repair to sections of trail. development of Grand Teton National Park as it joined the National Park Service system. Pre-existing Trails Archeological sites are present in the upper INTRODUCTION parts of Berry Creek drainage, thought to represent ―basecamps‖ occupied consistently over 8,000 years. Grand Teton National Park and its trail A notable pre-historic travel route traversed the system developed together during the early years of northern end of the Teton Range, from the west into National Park Service (NPS) administration. -

GRAND TETON National Park
Nationale Parken van Amerika @ www.ontdek-amerika.nl Last Update : 12 augustus 2015 GRAND TETON National Park KORTE OMSCHRIJVING Grootte : 1.255 vierkante kilometer Nationaal park sinds : 14 september 1950 Laagste punt : 1.935 meter (zuidgrens) Hoogste punt : 4.198 meter (piek van Grand Teton) De Grand Teton Mountains vormen een van de jongste bergketens van Noord-Amerika. Er zijn 12 pieken die hoger zijn dan 3650 meter; de hoogste is Grand Teton met een top van 4.198 meter. Het gebergte is ontstaan naast een breuklijn in de aarde; aan de westzijde van de breuklijn werd de aarde omhooggedrukt, aan de oostzijde zakte de aarde juist naar beneden. De 130 kilometer lange Teton Range heeft dan ook geen voorgebergte, de bergen rijzen direct naast de vloer van de vallei omhoog. Nog maar 15.000 jaar geleden hebben gletsjers de canyons tussen de bergpieken geslepen. De tongen van de gletsjers hebben ook de vallei bereikt, waardoor zich veel mooie meren hebben gevormd. De Jackson Hole vallei is 72 kilometer lang en 10 tot 20 kilometer breed, en er leven hier erg veel vogels en wilde dieren, waaronder zwarte beren, grizzlyberen, elanden, prairiewolven enzovoort. In het park worden erg veel activiteiten georganiseerd op het gebied van watersport, winter- sport, bergsport ed. Maar ook voor de toerist die vooral komt voor mooie uitkijkpunten en eenvoudige natuurwandelingen heeft het park veel te bieden BEREIKBAARHEID Het park ligt in het noordwesten van de staat Wyoming, 11 mijl onder Yellowstone National Park. Er zijn drie ingangen die aan hoofdwegen liggen, en er is één ingang die aan een kleine toegangsweg ligt. -

Spatial Structure of Melanism in Yellow-Bellied Marmots
Montopoli et al.: Spatial Structure of Melanism in Yellow-Bellied Marmots SPATIAL STRUCTURE OF MELANISM IN YELLOW-BELLIED MARMOTS GEORGE MONTOPOLI + NICK VISSER + MICHAEL CRONE UNIVERSITY OF WYOMING + LARAMIE + ABSTRACT incidence of melanism in the peppered moth Riston bistularia could be explained in terms of differential Melanism (black fur coloration) in the vulnerability to avian predators in a changing yellow-bellied marmot, Marmot a flaviventris, is environment. The normal light-colored phase of encountered uniquely in the Teton Range in peppered moths was highly cryptic on tree bark, and northwest Wyoming, This study is designed to melanistic phenotypes were rare in pre-industrial investigate whether the occurrence of melanism is England. With the large amounts of particulate associated with reduced predation due to high pollutants emitted by nineteenth century factories, human activity. Because overuse by humans can the tree bark on which the moths usually rested particularly stress the environment, the implications became darker, so that normal-phased moths were of this project are especially significant for Grand no longer cryptic. Consequently, melanistic Teton National Park where efforts are directed to phenotypes enjoyed the protection conferred by minimizing human impact. In addition, an crypticity, selection favored the melanistic allele, estimation technique to calculate the likelihood that and most moths were melanistic by the 1930's. The the melanistic allele is dominant is developed. peppered moth presents one of the clearest examples of short-term evolution in a free-ranging animal population, and of human influence on animal + INTRODUCTION coevolution. The yellow-bellied marmot in the Tetons The biology of melanism in the yellow of short-term evolution bellied marmot Marmot a flaviventris found in Grand may present another example influenced by humans. -

Structural Geology and History of the Buck Mountain Fault and Adjacent
Structural geology and history of the Buck Mountain fault and adjacent intra-range faults, Teton Range, Wyoming by Daniel Joseph Smith A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science in Earth Sciences Montana State University © Copyright by Daniel Joseph Smith (1991) Abstract: The Teton Range in northwestern Wyoming lies on the western margin of Laramide basement-involved deformation in the Rocky Mountain foreland. The study area is located in the central Teton Range and includes the high peaks of the range (the Cathedral Group) and several basement-involved deformation zones. The purpose of this study is to: 1) determine the geometry, kinematics and age of the Buck Mountain, Stewart and Static faults in the central Teton Range, and 2) to characterize post-Archean basement deformation in the Teton Range by examining mesoscopic and microscopic fabrics and alteration mineralogies in the study area. The north-striking Buck Mountain and northeast-striking Stewart faults exhibit brittle deformation including closespaced, unstable fractures and cataclasites. Slickenlines and drag folds indicate reverse dip-slip on the Buck Mountain fault. Both faults are interpreted to be Laramide on the basis of their basement-involved, contractional nature and brittle deformation. Static fault strikes northeast, dips steeply southeast and can be divided into two segments: 1) a southwest segment characterized by mylonitic rocks and exhibiting oriented chlorite, relict hornblende crystals, and rotated quartzo-feldspathic porphyroblasts (interpreted to be a Proterozoic phyllonite zone); and 2) a northeast segment that was reactivated by the Buck Mountain fault and exhibits brecciation and cataclasites. -
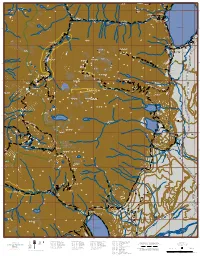
Grand Teton Loop Trail
-110.870 -110.860 -110.850 -110.840 -110.830 -110.820 -110.810 -110.800 -110.790 -110.780 -110.770 -110.760 -110.750 -110.740 -110.730 -110.720 Symmetry Spire Hang 0 in 0 g Rock of Ages 4 Cube Point 7, C 8 a , n 0 2 y on 0 0 Tr 8 0 , 9 0 0 7,0 00 8 r , T 9 e k ,800 0 a 8 0 L ,2 ny Ice Point 00 7 Jen 8 , 2 0 43.770 N 0 ,4 43.770 9, 00 o 7 2 r t Horse T h Storm Point ra F il 0 o 60 r 9, k 0 8,0 0 Inspiration Point 0 8 0 , Horse Tr Jct 7 0.75 r rk T C e d 0 Boat Dock - West n ca 7, 20 00 o s 4 a 9, y 0 n C 7,80 Ca ade 0.25 Forks of Cascade Canyon sc 0,000 a 1 scade C Hidden Falls Jenny Lake Tr 0 Ca Can 3.50 - L - yon T L H 0 GT r T o 0 ,2 G 9 Horse Tr Jct rs 0 e 8 Cascade Canyon T , r a 6 7,20 i 0 0 l 7, ,40 60 7 0 e Crk scad 8,000 Ca 00 H 7,8 o rs e T r a i 1.20 l 8,2 00 Jenny Lake 43.760 43.760 0 0 ,2 8 7 ,0 0 0 0 0 r 0 T , e 9 k 0 a 0 L 6 , ny 8 0 Jen Jenny Lake 40 8, H o Campground rs G e T 0 T L 0 r a 2 - , i l 7 J e n n Jenny Lake 0 0 y 6 , Ranger Station 9 L Valhalla Canyon a 0 k 0 e 7,6 T Boat Dock - East South Fork Cascade Canyon r 4.00 Jenny Lk Trailhead Horse Trail Jct 43.750 r h 43.750 0 T e t 0 k 4 a a d , 0.15 L P a 8 y o Moose Ponds Jct n e n 6,800 s R e U k J - r 0.15 Exum Guides i lt a u P ,800 M n 8 Mount Teewinot, 12,325 Valley Tr Jct to e Mount Owen, 12,928 T Moose Ponds Table Mountain ead M M ow 0 0 o e s P 0 11,6 0 0 o in ark 4 0 4 s , 0 0 A , 0 e p c 0 2 0 , ce 1 0 s 1 1 P u s 0 r 1 1 L 0 0 0 o , , 0 n 9 T ,2 d 2 0 1 s n 1 r 0.50 o T y East Face Rte n y a e l C ,600 l de 11 a a V c s - a L C 0 the Apex T 0 k 4 r , -

Grand Teton, 106–107 Best Trails, 5–6 Yellowstone, 40–41 Grand Teton, 134–136 Accommodations
13_769827 bindex.qxp 1/10/06 3:01 PM Page 238 Index See also Accommodations and Restaurant indexes below. GENERAL INDEX Back Basin Loop (Yellowstone), 53, 83 Access/entry points Backcountry Grand Teton, 106–107 best trails, 5–6 Yellowstone, 40–41 Grand Teton, 134–136 Accommodations. See also Accommo- planning a trip to, 37–39 dations Index Yellowstone, 90–98 best, 7, 8 Bechler Region, 95–96 Cody, 208–210 information, 91 Gardiner, 174–175 maps, 92–93 Grand Teton, 157–161 outfitters, 93 West Yellowstone, 170–172 permits, 23, 28, 92 Yellowstone, 73, 142–150 Shoshone Lake, 93–95 accessible, 34 Slough Creek Trail, 97–98 AdventureBus (Yellowstone), 79 Sportsman Lake Trail, 97 Adventures Beyond Yellowstone, 101 Thorofare area, 96–97 Adventure Sports (Moose), 137, 138, when to go, 92 180 Backcountry Adventure Snowmobile Aerial touring, Jackson, 185 Rental (West Yellowstone), 105 Airports, 29–30 Bakers Hole campground Alaska Basin (Grand Teton), 136 (Yellowstone), 152 Albright Visitor Center (Yellowstone), Bald eagle, 235 41, 50, 58 Ballooning, Jackson, 185 Altitude sickness, 36 Barker-Ewing Float Trips (Grand American dipper, 237 Teton), 140 American white pelicans, 236 Bears (black and grizzly), 16, 86, 88, Amphitheater Lake Trail (Grand 89, 228–230 Teton), 130 encounters with, 36–37 Anemone Geyser (Yellowstone), Grizzly and Wolf Discovery Center 75, 89 (West Yellowstone), 169 Angel Terrace (Yellowstone), 58 Beartooth Highway, 23 Antelope Flats Road (Grand Teton), Beaverdam Creek (Yellowstone), 97 120, 136 Beaver Lake Picnic Area Art galleries, -

Wyoming Road Trip by the Mile Marker
Available at Amazon.com BROOK BESSER WYOMING ROAD TRIP BY THE MILE MARKER Travel guide to Yellowstone/Grand Teton National Park, Devils Tower, Oregon/Mormon Trail, badlands, petroglyphs, waterfalls, camping, hiking, tourism and more... Brook Besser NightBlaze Books Golden, Colorado Copyright © 2010 by Brook Besser All rights reserved. No part of this book may be reproduced or transmitted in any form by any means, electronic or mechanical, including photocopying and scanning, or stored in any information storage and retrieval system, without the express consent of the author. ISBN 978-0-9844093-0-3 Library of Congress Control Number: 2010923145 Manufactured in the United States of America WARNING! NightBlaze Books and the author assume no responsibility or liability for any damages, losses, accidents, or injuries incurred by readers who visit the attractions or engage in the activities described in this book. It is the reader’s responsibility to be aware of all risks and take the necessary precautions to handle those risks. The! suggestions and "Cool Ratings" in this book are strictly the author's opinion, and expressed to help you make a decision on whether to visit an attraction. You are responsible for making your own judgment on the worthiness of an attraction. Special thanks to the following people: My wife Mianne who accompanied me for 5 weeks in Wyoming and supported my endless hours working on this book; my brother Brant and my daughter Brianna who each spent a week out on the road with me; my brother Brett who helped with my book summaries; my sister-in-law Sue who proofed many pages of the book; and the rest of my family for their support. -

1988 Yellowstone Fires! 20 Years After FREE
MOUNTAINMOUNTAIN COUNTRY COUNTRY 2008 Traveler’s Guide to Grand Teton & Yellowstone Vacation Adventures Boating • Hiking • Climbing Biking • Rodeo • Fishing Mountain Towns National Parks Area Map Wildlife 1988 Yellowstone Fires! 20 Years After FREE Jewelry Originals 32 YEARS OF INSIRATION AT 6,000 FT. Gaslight Alley • Downtown Jackson Hole • 125 N.Cache www.DanShelley.com • [email protected] • 307.733.2259 ALL DESIGNS COPYRIGHTED Jackson Hole’s Best Real Estate Opportunities Hotel Terra Jackson Hole 20 Winger Circle, Teton Springs, Idaho Ski in/ski out property in Teton Village. Combining supreme Recently completed 4 bedroom, 4.5 bathroom home with nearly luxury and environmental sustainability, Hotel Terra is a 72- 4,000 sq. ft. of living space and a 3 car garage. This home has amaz- room, slope side eco-boutique hotel, offering two restaurants, ing views from every window of either the golf course or the Teton the Jackson Hole Tree house ski/snowboard rental shop and Range. Take advantage of all the amenities this four season resort “Chill” rooftop spa and hot tub. Phase II scheduled opening in community has to offer including club house, restaurants, tennis, 2009. Pricing from $1,395,000 to $2,600,000. pools, golf and skiing. Offered at $1,350,000 307.733.4159 800.543.6328 are qu S n Hwy 22 w o T S Albertson’s ou 89 Hwy th P ark Loop Smith’s H We’re a Jackson o High School Rd b a Hole MUST-SEE! c k Try free samples "UFFALO%LK in our factory 3TEAK0ACK store on Hwy 89 100% Natural .ATURAL at Smith’s Plaza. -

Seasonal and Altitudinal Variation in Pollinator Communities in G 5
Dillon: Seasonal and Altitudinal Variation in Pollinator Communities in G 5 SEASONAL AND ALTITUDINAL VARIATION IN POLLINATOR COMMUNITIES IN GRAND TETON NATIONAL PARK MICHAEL E. DILLON DEPARTMENT OF ZOOLOGY AND PHYSIOLOGY AND PROGRAM IN ECOLOGY UNIVERSITY OF WYOMING LARAMIE ABSTRACT over the last 20-30 years. These pollinator declines are alarming not only because of their effects on Native pollinators are in decline across the agriculture (Berenbaum et al. 2007) and therefore globe, likely due to a combination of habitat loss, human health (Eilers et al. 2011) but also because of pesticides, invasive species and changing climate. their potentially substantial and far-reaching indirect Determining the independent effects of climate on effects on ecosystem services (over 85% of flowering pollinators has been difficult in part because we lack plants depend on insect pollination; Ollerton et al. studies of pollinator populations in largely 2011). undisturbed areas. Early spring and alpine pollinators are most likely to be affected by changing climate. Pollinator declines are undoubtedly tied to Using a standardized sampling protocol, I measured loss of habitat associated with changes in landscape relative abundance of major pollinator groups (flies, use (Cameron et al. 2011), to nontarget effects of beetles, bees, wasps, and butterflies) from early over- and misuse of common herbicides and spring to late summer at sites ranging from 2100 to insecticides (Henry et al. 2012, Whitehorn et al. 3300 m elevation. Flies were most abundant in early 2012), and to pathogen spread from introduced spring and at high elevations. Bees were abundant species (Stout and Morales 2009). However, the throughout the season and across all elevations. -

GRAND TETON NATIONAL PARK Grand Teton NATIONAL PARK
GRAND TETON NATIONAL PARK Grand Teton NATIONAL PARK WYOMING OPEN ALL YEAR Contents History of the Region 6 Dude Ranches . 21 Geographic Features 8 Administration 22 Teton Range 8 How To Reach the Park 22 Jackson Hole 9 By Automobile 22 The Work of Glaciers 10 By Railroad and Bus 24 Trails 12 By Airplane 24 Mountain Climbing 14 Points of Interest Along the Way .... 24 Wildlife 15 Trees and Plants 19 Accommodations and Expenses .... 25 Naturalist Service 21 Bibliography 28 Fishing 21 Government Publications 30 Swimming 21 National Parks in Brief 31 Hunting 21 Rules and Regulations 32 Events OF HISTORICAL IMPORTANCE I 879 Thomas Moran painted the Teton Range. I 884 The first settlers entered Jackson Hole. 1807—8 Discovery of the Tetons by John Colter. I 897 Teton Forest Reserve created. 1811 The Astorians crossed Teton Pass. 1898 The first major Teton peaks scaled (Buck Mountain and Grand Teton). 1810—45 "The Fur Era" in the Rocky Mountains, which reached its height between 1825 and 1840. I909 The Upper Gros Venrre landslide. 1829 Capt. William Sublette named Jackson Hole after his partner in the 1925 The Lower Gros Ventre landslide. fur trade, David Jackson. 1927 The Gros Ventre flood. I 832 Rendezvous of the fur trappers in Pierre's Hole; the Battle of Pierre's 1929 Grand Teton National Park created and dedicated. Hole. 1930 The last major Teton peaks scaled (Nez Perce and Mount Owen). 1835 Rev. Samuel Parker conducted the first Protestant service in the Rocky Mountains a few miles south of the Tetons. I 843 Michaud attempted an ascent of the Grand Teton. -

Copyrighted Material
INDEX See also Accommodations and Restaurant indexes, below. GENERAL INDEX Back Basin Loop (Yellowstone), 55, 83 Academic trips, 36–37 Backcountry, 5–6, 40 Access/entry points Grand Teton, 130–132 Grand Teton, 105, 108 permits, 39, 91–92, 130 Yellowstone, 43, 46 Yellowstone, 91–97 Accessibility, 31–32 Backpacking for beginners, 41 Accommodations, 38–39. See also Backroads, 37 Camping and campgrounds Bakers Hole campground (West best, 7, 8 Yellowstone), 146 Cody, 200–203 Ballooning, Jackson, 178 env ironmentally-friendly, 35 Barker-Ewing (Jackson), 175 Gardiner, 168–169 Barker-Ewing Float Trips (Jackson), Grand Teton, 151–155 136 Jackson and environs, 180–188 Bears, 15, 16, 30–31, 217–219 West Yellowstone, 164–166 Beartooth Highway, 21 Yellowstone National Park, Beaver Lake Picnic Area 138–144 (Yellowstone), 56 AdventureBus, 37, 79 Beaver Ponds Loop Trail Adventure Sports (Moose), 133, (Yellowstone), 60, 84 134, 173 Bechler Meadows Trail Adventure trips, 37 (Yellowstone), 95–96 Aerial touring, Jackson, 178 Bechler Meadows Trails Alaska Basin (Grand Teton), 132 (Yellowstone), 96 Albright Visitor Center (Yellow- The Bechler Region (Yellowstone), stone), 46, 52, 58–59 94–95 Altitude sickness, 28 Bechler River Trail (Yellowstone), Amphitheater Lake Trail (Grand 95, 96 Teton), 127 Big Wild Adventures, 93 Anemone Geyser (Yellowstone), 74 Biking, 48, 98, 132–133, 196 Angel Terrace (Yellowstone), 59 Billings (Montana), 25 Antelope Flats (Grand Teton), Birds, 223–225 132–133 Biscuit Basin (Yellowstone), 73, 76 AntelopeCOPYRIGHTED Flats Road (Grand