Evolution of Colour Vision in Vertebrates
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Visual Purple and the Photopic Luminosity Curve
Br J Ophthalmol: first published as 10.1136/bjo.32.11.793 on 1 November 1948. Downloaded from THE BRITISH JOURNAL OF OPHTHALMOLOGY NOVEMBER, 1948 by copyright. COMMUNICATIONS VISUAL PURPLE AND THE PHOTOPIC LUMINOSITY- CURVE* BY H. J. A. DARTNALL http://bjo.bmj.com/ FROM THE VISION RESEARCH UNIT, 'MEDICAL RESEARCH COUNCIL, INSTITUTE OF OPHTHALMOLOGY, LONDON Introduction THE precise relationship which has been established between the scotopic luminosity curve a'nd visual purple (Dartnall & Goodeve, 1937; Wald, 19138) leads one to expect that the photopic luminositv on September 26, 2021 by guest. Protected curve is'similarly related to some " photopic " pigment or pig- ments. This latter hypothesis has given rise to considerable speculation, particularly since the evidence for the existence of retinal pigments, other than the various forms of visual purple, is not unequivocal. In any case there is no evidence for such additional pigments in the human retina. Visual purple mediates scotopic- vision by virtue of the photo- chemical changes it undergoes on exposure to light. The rate of * Received for Publication, July 12, 1948. Br J Ophthalmol: first published as 10.1136/bjo.32.11.793 on 1 November 1948. Downloaded from 794 H. J. A. DARTNALL any photochemical change is gQverned by the value of the product y Ia where y is the quantqm efficiency of the process and Ia the intensity of the absorbed light expressed in quanta per second. Since the eye in the photopic condition is much less sensitive than in the scotopic state it follows that the value of y la must be correspondingly smaller for the photopic process. -

The Purkinje Rod-Cone Shift As a Function of Luminance and Retinal Eccentricity
Vision Research 42 (2002) 2485–2491 www.elsevier.com/locate/visres Rapid communication The Purkinje rod-cone shift as a function of luminance and retinal eccentricity Stuart Anstis * Department of Psychology, University of California, San Diego, 9500 Gilman Drive, La Jolla, CA 92093-0109, USA Received 14 September 2000; received in revised form 25 June 2002 Abstract In the Purkinje shift, the dark adapted eye becomes more sensitive to blue than to red as the retinal rods take over from the cones. A striking demonstration of the Purkinje shift, suitable for classroom use, is described in which a small change in viewing distance can reverse the perceived direction of a rotating annulus. We measured this shift with a minimum-motion stimulus (Anstis & Cavanagh, Color Vision: Physiology & Psychophysics, Academic Press, London, 1983) that converts apparent lightness of blue versus red into apparent motion. We filled an iso-eccentric annulus with radial red/blue sectors, and arranged that if the blue sectors looked darker (lighter) than the red sectors, the annulus would appear to rotate to the left (right). At equiluminance the motion appeared to vanish. Our observers established these motion null points while viewing the pattern at various retinal eccentricities through various neutral density filters. Results: The luminous efficiency of blue (relative to red) increased linearly with eccentricity at all adaptation levels, and the more the dark-adaptation, the steeper the slope of the eccentricity function. Thus blue sensitivity was a linear function of eccentricity and an exponential function of filter factor. Blue sensitivity increased linearly with eccentricity, and each additional log10 unit of dark adaptation changed the slope threefold. -

TETRACHROMATIC COLOR VISION Kimberly A. Jameson Prepared For
TETRACHROMATIC COLOR VISION Kimberly A. Jameson prepared for The Oxford Companion to Consciousness. Wilken, P., Bayne, T. & Cleeremans, A. (Ed.s). Oxford University Press: Oxford. To appear in 2007. The term“tetrachromacy” describes the physiological possession of four different classes of simultaneously functioning retinal photopigments (also called “weak tetrachromacy”). From an empirical standpoint, tetrachromatic color vision (or “strong tetrachromacy”) additionally requires demonstrating that mixtures of four independent appropriately chosen primary lights will simulate all distinc- tions in appearance possible in visible color space. Independence of the primary lights implies that no mixtures of any subset of these lights (or their intensity variants) will produce an identical match to any combination of mixtures of the remaining lights. By comparison, trichromacy empirically requires only three primaries to simulate all visible colors. Established theory states that normal color vision humans are trichromats (as, primarily, are Old World monkeys and apes). The first element of trichro- macy is the output from three simultaneously functioning retinal cone classes: Short-, Medium-, and Long- Wavelenth Sensitive (SWS, MWS, & LWS) cones. Three cone classes alone do not establish a trichromat color code, however. A postreceptoral code for three categories of signal is also needed. A standard vision science assumption is that the postreceptoral recoding of cone outputs initiates the neural trivariant (or trichromatic) property of human color percep- tion, and the need for only three primary lights to match any test light. ANIMAL TETRACHROMACY: Tetrachromacy is an early vertebrate characteristic, existing in fish and reptiles, and is evolutionarily more ancient than primate trichromacy. Essentially all diu- ral birds have four retinal cone types (two SWS classes, plus a MWS and a LWS class) which neurally produce four dimensional color experience, or tetrachro- matic color vision. -
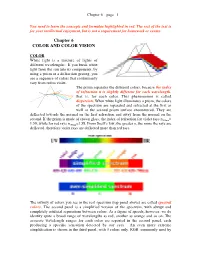
Chapter 6 COLOR and COLOR VISION
Chapter 6 – page 1 You need to learn the concepts and formulae highlighted in red. The rest of the text is for your intellectual enjoyment, but is not a requirement for homework or exams. Chapter 6 COLOR AND COLOR VISION COLOR White light is a mixture of lights of different wavelengths. If you break white light from the sun into its components, by using a prism or a diffraction grating, you see a sequence of colors that continuously vary from red to violet. The prism separates the different colors, because the index of refraction n is slightly different for each wavelength, that is, for each color. This phenomenon is called dispersion. When white light illuminates a prism, the colors of the spectrum are separated and refracted at the first as well as the second prism surface encountered. They are deflected towards the normal on the first refraction and away from the normal on the second. If the prism is made of crown glass, the index of refraction for violet rays n400nm= 1.59, while for red rays n700nm=1.58. From Snell’s law, the greater n, the more the rays are deflected, therefore violet rays are deflected more than red rays. The infinity of colors you see in the real spectrum (top panel above) are called spectral colors. The second panel is a simplified version of the spectrum, with abrupt and completely artificial separations between colors. As a figure of speech, however, we do identify quite a broad range of wavelengths as red, another as orange and so on. -

Adjustment of Lighting Parameters from Photopic to Mesopic Values in Outdoor Lighting Installations Strategy and Associated Evaluation of Variation in Energy Needs
sustainability Article Adjustment of Lighting Parameters from Photopic to Mesopic Values in Outdoor Lighting Installations Strategy and Associated Evaluation of Variation in Energy Needs Enrique Navarrete-de Galvez 1 , Alfonso Gago-Calderon 1,* , Luz Garcia-Ceballos 2, Miguel Angel Contreras-Lopez 2 and Jose Ramon Andres-Diaz 1 1 Proyectos de Ingeniería, Departamento de Expresión Gráfica Diseño y Proyectos, Universidad de Málaga, 29071 Málaga, Spain; [email protected] (E.N.-d.G.); [email protected] (J.R.A.-D.) 2 Expresión Gráfica en la Ingeniería, Departamento de Expresión Gráfica Diseño y Proyectos, Universidad de Málaga, 29071 Málaga, Spain; [email protected] (L.G.-C.); [email protected] (M.A.C.-L.) * Correspondence: [email protected]; Tel.: +34-951-952-268 Abstract: The sensitivity of the human eye varies with the different lighting conditions to which it is exposed. The cone photoreceptors perceive the color and work for illuminance conditions greater than 3.00 cd/m2 (photopic vision). Below 0.01 cd/m2, the rods are the cells that assume this function (scotopic vision). Both types of photoreceptors work coordinately in the interval between these values (mesopic vision). Each mechanism generates a different spectral sensibility. In this work, Citation: Navarrete-de Galvez, E.; the emission spectra of common sources in present public lighting installations are analyzed and Gago-Calderon, A.; Garcia-Ceballos, their normative photopic values translated to the corresponding mesopic condition, which more L.; Contreras-Lopez, M.A.; faithfully represents the vision mechanism of our eyes in these conditions. Based on a common Andres-Diaz, J.R. Adjustment of street urban configuration (ME6), we generated a large set of simulations to determine the ideal light Lighting Parameters from Photopic to point setup configuration (luminance and light point height vs. -

Color Vision Test for Dichromatic and Trichromatic Macaque Monkeys
Journal of Vision (2013) 13(13):1, 1–15 http://www.journalofvision.org/content/13/13/1 1 Color vision test for dichromatic and trichromatic macaque monkeys National Institute for Physiological Sciences, Okazaki, Aichi, Japan Electronics-Inspired Interdisciplinary Research Institute, Toyohashi University of Technology, Toyohashi, Aichi, # Kowa Koida* Japan $ National Institute for Physiological Sciences, Okazaki, Aichi, Japan University for Advanced Studies (SOKENDAI), Okazaki, # Isao Yokoi* Aichi, Japan $ National Institute for Physiological Sciences, Okazaki, Aichi, Japan University for Advanced Studies (SOKENDAI), Okazaki, # Gouki Okazawa Aichi, Japan $ Faculty of Rehabilitation, Chubu Gakuin University, Seki, # Akichika Mikami Gifu, Japan $ Department of Biology, Bogor Agricultural University, # Kanthi Arum Widayati Bogor, Indonesia $ Primate Research Institute, Kyoto University, Inuyama, # Shigehiro Miyachi Aichi, Japan $ National Institute for Physiological Sciences, Okazaki, Aichi, Japan University for Advanced Studies (SOKENDAI), Okazaki, # Hidehiko Komatsu Aichi, Japan $ Dichromacy is a color vision defect in which one of the identified dichromatic macaques (Macaca fascicularis) three cone photoreceptors is absent. Individuals with with that of normal trichromatic macaques. In the color dichromacy are called dichromats (or sometimes ‘‘color- discrimination test, dichromats could not discriminate blind’’), and their color discrimination performance has colors along the protanopic confusion line, though contributed significantly -

Human Trichromacy Revisited PNAS PLUS
Human trichromacy revisited PNAS PLUS Hiroshi Horiguchia,b,1, Jonathan Winawera, Robert F. Doughertyc, and Brian A. Wandella,c aPsychology Department, Stanford University, Stanford, CA 94305; bDepartment of Ophthalmology, School of Medicine, Jikei University, Tokyo 105-8461, Japan; and cStanford Center for Cognitive and Neurobiological Imaging, Stanford, CA 94305 AUTHOR SUMMARY The history of trichromatic color As predicted by the classic tri- theory spans nearly 300 y, be- msARC Fovea Periphery chromatic model, subjects fail to ’ Opticks (%) ginning with Newton s 10 see cone-silent stimuli presented (1). Thomas Young’s Royal So- Visible in the fovea. In contrast, subjects Not ciety Lecture (2) explained how 0 Not do see cone-silent stimuli pre- Visible Visible Visible Visible color perception is initiated by S1 sented to the peripheral visual S2 fi the absorption of light in three Cone-silent −10 eld. In extensive computational S3 fi types of human light receptors, −10 0 10 −10 0 10 (%) modeling, we could nd no plau- the cones. The theory of tri- S-cone S-cone sible pigment density adjustment chromacy is the foundation of (including the lens or photopig- modern color technologies (3). Fig. P1. Built and found. (A) An apparatus using six primary lights was ment densities) that explains the built for the experiment. Mean luminance through the eyepiece was measurements in the periphery For more than a century, the 2 absorption of light in the retina 2,060 cd/m .(B) Graphs represent detection thresholds for stimuli that based on a model that includes was thought to occur only in the have some S-cone contrast and cone-silent contrast, but no L- or M- only three cone photopigments. -

Colour Vision Deficiency
Eye (2010) 24, 747–755 & 2010 Macmillan Publishers Limited All rights reserved 0950-222X/10 $32.00 www.nature.com/eye Colour vision MP Simunovic REVIEW deficiency Abstract effective "treatment" of colour vision deficiency: whilst it has been suggested that tinted lenses Colour vision deficiency is one of the could offer a means of enabling those with commonest disorders of vision and can be colour vision deficiency to make spectral divided into congenital and acquired forms. discriminations that would normally elude Congenital colour vision deficiency affects as them, clinical trials of such lenses have been many as 8% of males and 0.5% of femalesFthe largely disappointing. Recent developments in difference in prevalence reflects the fact that molecular genetics have enabled us to not only the commonest forms of congenital colour understand more completely the genetic basis of vision deficiency are inherited in an X-linked colour vision deficiency, they have opened the recessive manner. Until relatively recently, our possibility of gene therapy. The application of understanding of the pathophysiological basis gene therapy to animal models of colour vision of colour vision deficiency largely rested on deficiency has shown dramatic results; behavioural data; however, modern molecular furthermore, it has provided interesting insights genetic techniques have helped to elucidate its into the plasticity of the visual system with mechanisms. respect to extracting information about the The current management of congenital spectral composition of the visual scene. colour vision deficiency lies chiefly in appropriate counselling (including career counselling). Although visual aids may Materials and methods be of benefit to those with colour vision deficiency when performing certain tasks, the This article was prepared by performing a evidence suggests that they do not enable primary search of Pubmed for articles on wearers to obtain normal colour ‘colo(u)r vision deficiency’ and ‘colo(u)r discrimination. -

1 Human Color Vision
CAMC01 9/30/04 3:13 PM Page 1 1 Human Color Vision Color appearance models aim to extend basic colorimetry to the level of speci- fying the perceived color of stimuli in a wide variety of viewing conditions. To fully appreciate the formulation, implementation, and application of color appearance models, several fundamental topics in color science must first be understood. These are the topics of the first few chapters of this book. Since color appearance represents several of the dimensions of our visual experience, any system designed to predict correlates to these experiences must be based, to some degree, on the form and function of the human visual system. All of the color appearance models described in this book are derived with human visual function in mind. It becomes much simpler to understand the formulations of the various models if the basic anatomy, physiology, and performance of the visual system is understood. Thus, this book begins with a treatment of the human visual system. As necessitated by the limited scope available in a single chapter, this treatment of the visual system is an overview of the topics most important for an appreciation of color appearance modeling. The field of vision science is immense and fascinating. Readers are encouraged to explore the liter- ature and the many useful texts on human vision in order to gain further insight and details. Of particular note are the review paper on the mechan- isms of color vision by Lennie and D’Zmura (1988), the text on human color vision by Kaiser and Boynton (1996), the more general text on the founda- tions of vision by Wandell (1995), the comprehensive treatment by Palmer (1999), and edited collections on color vision by Backhaus et al. -

Color Vision Deficiency
Color Vision Deficiency What is color vision deficiency? Color vision deficiency is called “color blindness” by mistake. Actually, the term describes a number of different problems people have with color vision. Abnormal color vision may vary from not being able to tell certain colors apart to not being able to identify any color. Whom does color vision deficiency affect? An estimated 8% of males and fewer than 1% of females have color vision problems. Most color vision problems run in families and are inherited and present at birth. A child inherits a color vision deficiency by receiving a faulty color vision gene from a parent. Abnormal color vision is found in a recessive gene on the X chromosome. Men are born with just one X and one Y chromosome. However, women have two X chromosomes. Because of this, women can sometimes overcome the faulty gene with their second normal X chromosome. Men, unfortunately, do not have a second X chromosome to help compensate for the faulty color vision gene. Heredity does not cause all color vision problems. One common problem happens from the normal aging of the eye’s lens. The lens is clear at birth, but the aging process causes it to darken and yellow. Older adults may have problems identifying certain dark colors, particularly blues. Certain medications as well as inherited or acquired retinal and optic nerve disease, may also affect normal color vision. Who should be tested for color deficiency? Any child who is having difficulty in school should be checked for possible visual problems including color vision impairment. -

Reptilian Eyes and Orbital Structures
REPTILIAN EYES AND ORBITAL STRUCTURES Jeanette Wyneken, PhD Florida Atlantic University, Dept. of Biological Sciences, 777 Glades Road, Boca Raton, FL 33431 USA ABSTRACT The anatomy of the reptilian eye is similar across species, generally, but the eyes of various taxa differ in details. The eyeball is formed of layers and has three chambers. Pupil shape differs among reptilian taxa with behavior. The retina’s photosensitive cells (rods and cones or all- cones) transmit signals to the optic nerve, which is an extension of the brain. Retinal sensitivity is increased by foveae in lizards and tuataras, while turtles and snakes have areas. The lens is soft and is shaped for accommodation by the ciliary muscles (lizards, turtles and crocodilians) or movement (snakes). All reptiles have eyelids, however some lizards have partially fused lids, while other lizards and all snakes have fused clear lids, the spectacle. Ocular glands lubricate the cornea in all species. Movements of the eyes are critical to preventing photoreceptor fatigue and loss of image recognition. INTRODUCTION TO THE REPTILIAN EYE Reptilian eyes are anatomically similar to those of other vertebrates in that the eyeball (= globe) is formed of layers, filled with fluid, and has a lens that focuses light on a retina. The eye is structured as a series of chambers. The anterior chamber is the fluid-filled space inside the eye between the iris and the cornea's innermost surface; the posterior chamber is a small space directly posterior to the iris, anterior to the lens, and bordered by the ciliary body or ciliary muscles. The anterior and posterior chambers are filled with aqueous humour. -

17-2021 CAMI Pilot Vision Brochure
Visual Scanning with regular eye examinations and post surgically with phoria results. A pilot who has such a condition could progress considered for medical certification through special issuance with Some images used from The Federal Aviation Administration. monofocal lenses when they meet vision standards without to seeing double (tropia) should they be exposed to hypoxia or a satisfactory adaption period, complete evaluation by an eye Helicopter Flying Handbook. Oklahoma City, Ok: US Department The probability of spotting a potential collision threat complications. Multifocal lenses require a brief waiting certain medications. specialist, satisfactory visual acuity corrected to 20/20 or better by of Transportation; 2012; 13-1. Publication FAA-H-8083. Available increases with the time spent looking outside, but certain period. The visual effects of cataracts can be successfully lenses of no greater power than ±3.5 diopters spherical equivalent, at: https://www.faa.gov/regulations_policies/handbooks_manuals/ techniques may be used to increase the effectiveness of treated with a 90% improvement in visual function for most One prism diopter of hyperphoria, six prism diopters of and by passing an FAA medical flight test (MFT). aviation/helicopter_flying_handbook/. Accessed September 28, 2017. the scan time. Effective scanning is accomplished with a patients. Regardless of vision correction to 20/20, cataracts esophoria, and six prism diopters of exophoria represent series of short, regularly-spaced eye movements that bring pose a significant risk to flight safety. FAA phoria (deviation of the eye) standards that may not be A Word about Contact Lenses successive areas of the sky into the central visual field. Each exceeded.