A Revision of Epithema (Gesneriaceae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Further Miscellaneous Species of Cyrtandra in Borneo
E D I N B U R G H J O U R N A L O F B O T A N Y 63 (2&3): 209–229 (2006) 209 doi:10.1017/S0960428606000564 E Trustees of the Royal Botanic Garden Edinburgh (2006) Issued 30 November 2006 OLD WORLD GESNERIACEAE XII: FURTHER MISCELLANEOUS SPECIES OF CYRTANDRA IN BORNEO O. M. HILLIARD &B.L.BURTT Nineteen miscellaneous species of Bornean Cyrtandra are dealt with. Cyrtandra atrichoides, C. congestiflora, C. crockerella, C. dulitiana, C. kanae, C. libauensis, C. plicata, C. vaginata and C. disparoides subsp. inconspicua are newly described. Descriptions and discussion are provided for C. erythrotricha and C. poulsenii, originally published with diagnoses only. Cyrtandra axillaris, C. longicarpa and C. microcarpa are also described, while C. borneensis, C. dajakorum, C. glomeruliflora, C. latens and C. prolata are reduced to synonymy. Keywords. Borneo, Cyrtandra, Gesneriaceae, new species. I NTRODUCTION A good many species of Cyrtandra in Borneo still remain undescribed; some available specimens are known to us only in the sterile state or are otherwise inadequate to typify a name. In this paper eight species and one subspecies are newly described. Burtt (1996) published new species with diagnoses only: C. erythrotricha B.L.Burtt and C. poulsenii B.L.Burtt are now fully described while C. glomeruliflora B.L.Burtt is reduced to synonymy under C. poulsenii. Full descriptions of C. axillaris C.B.Clarke and C. microcarpa C.B.Clarke are also given for the first time with the reduction of C. latens C.B.Clarke and C. dajakorum Kraenzl. -

December 2019/January 2020 Explore Norway with the Experts
DECEMBER 2019/JANUARY 2020 Explore Norway with the experts M11478 HUR Travel Bulletin FPx2 ad A4 v6.indd 1 14/11/2019 10:40 New Expedition Cruises Norway ––– With more than 125 years’ experience exploring Norway, this coastline is our home. Join us on our new Expedition Cruises and we’ll take you on an immersive, authentic voyage to the heart of Norway. Our year-round voyages on our new hybrid-powered expedition eet include nine handpicked, exclusive ports, and a passionate team of expedition experts to help you explore this extraordinary coastline. WHY Summer (May - September): HURTIGRUTEN? Fjords and Midnight Sun Expedition Norway In Handpicked itinerary Winter (October - February): Longer port stays Arctic Winter and Northern a Nutshell Included daily activities and a wide Lights Expedition range of optional excursions Spring (March - April): Each ship features a Science Center one, two or three day White Winter Expedition TRIPS oslo to bergen where you can interact with our via flåm and voss Expedition Team New Expedition Fleet Three onboard restaurants serving MS Otto Sverdrup For an immersive Norway delicious cuisine that refl ects our MS Eirik Raude add on coastal experience Norwegian heritage MS Maud Save AU$1,000 on selected 2021/22 departures go to hurtigruten.com.au call 1300 322 062 EMAIL [email protected] M11478 HUR Travel Bulletin FPx2 ad A4 v6.indd 2 14/11/2019 10:40 DEC 2019/JAN 2020 AIRLINE UPDATE USA SOLO TRAVELLERS LUXURY AUSTRALIA WINNERS AND LOSERS OF 2019 18 Part 2 Welcome to Sydney Explorer Dream Explorer -

On the Crabs of the Family Ocypodidae in the Collection of the Raffles Museum M. W. F. TWEEDIE
Reprint from Bulletin of the Raffles Museum, Singapore, Straits Settlements, No. IS, August 1937 On the Crabs of the family Ocypodidae in the collection of the Raffles Museum hy M. W. F. TWEEDIE M. W. F. TWEEDIE On the Crabs of the Family Ocypodidae in the Collection of the Raffles Museum By M. W. F. TwEEDiE, M.A. The material described in this paper has been collected for the most part during the last four years, mainly in mangrove swamps around Singapore Island and at a few localities on the east and west coasts of the Malay Peninsula. The greater part of the paper and most of the figures were prepared at the British Museum (Natural History) during August and September, 1936, and my grateful acknowledgments are due to the Director for permission to work there and for facilities provided, and particularly to Dr. Isabella Gordon for her unfailing help and encouragement. I wish also to express my thanks to the Directorates of the Zoological Museums at Leiden and Amsterdam for permission to examine types, and for the helpfulness and courtesy with which I was received by the members of the staffs of these museums. Finally acknowledgments are due to Prof. Dr. H. Balss, Dr. B. N. Chopra and Dr. C. J. Shen for their kindness in comparing specimens with types and authentic specimens in their respective institutions. The mode adopted for collecting the material may be of interest to collectors of Crustacea, and possibly other invertebrate groups, in the tropics. It was found that if crabs, especially Grapsidse and Ocypodidse, are put straight into alcohol, they tend to die slowly and in their struggles to shed their limbs and damage each other, so that often less than 10% of the collection survive as perfect specimens. -

Polygalaceae) from Borneo
Gardens' Bulletin Singapore 57 (2005) 47–61 47 New Taxa and Taxonomic Status in Xanthophyllum Roxb. (Polygalaceae) from Borneo W.J.J.O. DE WILDE AND BRIGITTA E.E. DUYFJES National Herbarium of the Netherlands, Leiden Branch P.O. Box 9514, 2300 RA Leiden, The Netherlands Abstract Thirteen new taxa or taxa with a new status in Xanthophyllum (Polygalaceae) from Borneo are described. The ten new species described in this paper are: X. bicolor W.J. de Wilde & Duyfjes, X. brachystachyum W.J. de Wilde & Duyfjes, X. crassum W.J. de Wilde & Duyfjes, X. inflatum W.J. de Wilde & Duyfjes, X. ionanthum W.J. de Wilde & Duyfjes, X. longum W.J. de Wilde & Duyfjes, X. nitidum W.J. de Wilde & Duyfjes, X. pachycarpon W.J. de Wilde & Duyfjes, X. rectum W.J. de Wilde & Duyfjes and X. rheophilum W.J. de Wilde & Duyfjes, and the new variety is X. griffithii A.W. Benn var. papillosum W.J. de Wilde & Duyfjes. New taxonomic status has been accorded to X. adenotus Miq. var. arsatii (C.E.C. Fisch.) W.J. de Wilde & Duyfjes and X. lineare (Meijden) W.J. de Wilde & Duyfjes. Introduction During the study of Xanthophyllum carried out in the BO, KEP, L, SAN, SAR and SING herbaria for the account of Polygalaceae in the Tree Flora of Sabah and Sarawak, several new taxa were defined. Their taxonomic position within the more than 50 species of Xanthophyllum recognised in Sabah and Sarawak will be clarified in the treatment of the family in the forthcoming volume of the Tree Flora of Sabah and Sarawak series. -

Report on Biodiversity and Tropical Forests in Indonesia
Report on Biodiversity and Tropical Forests in Indonesia Submitted in accordance with Foreign Assistance Act Sections 118/119 February 20, 2004 Prepared for USAID/Indonesia Jl. Medan Merdeka Selatan No. 3-5 Jakarta 10110 Indonesia Prepared by Steve Rhee, M.E.Sc. Darrell Kitchener, Ph.D. Tim Brown, Ph.D. Reed Merrill, M.Sc. Russ Dilts, Ph.D. Stacey Tighe, Ph.D. Table of Contents Table of Contents............................................................................................................................. i List of Tables .................................................................................................................................. v List of Figures............................................................................................................................... vii Acronyms....................................................................................................................................... ix Executive Summary.................................................................................................................... xvii 1. Introduction............................................................................................................................1- 1 2. Legislative and Institutional Structure Affecting Biological Resources...............................2 - 1 2.1 Government of Indonesia................................................................................................2 - 2 2.1.1 Legislative Basis for Protection and Management of Biodiversity and -

A Taxonomic Revision of Araceae Tribe Potheae (Pothos, Pothoidium and Pedicellarum) for Malesia, Australia and the Tropical Western Pacific
449 A taxonomic revision of Araceae tribe Potheae (Pothos, Pothoidium and Pedicellarum) for Malesia, Australia and the tropical Western Pacific P.C. Boyce and A. Hay Abstract Boyce, P.C. 1 and Hay, A. 2 (1Herbarium, Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3AE, U.K. and Department of Agricultural Botany, School of Plant Sciences, The University of Reading, Whiteknights, P.O. Box 221, Reading, RS6 6AS, U.K.; 2Royal Botanic Gardens, Mrs Macquarie’s Road, Sydney, NSW 2000, Australia) 2001. A taxonomic revision of Araceae tribe Potheae (Pothos, Pothoidium and Pedicellarum) for Malesia, Australia and the tropical Western Pacific. Telopea 9(3): 449–571. A regional revision of the three genera comprising tribe Potheae (Araceae: Pothoideae) is presented, largely as a precursor to the account for Flora Malesiana; 46 species are recognized (Pothos 44, Pothoidium 1, Pedicellarum 1) of which three Pothos (P. laurifolius, P. oliganthus and P. volans) are newly described, one (P. longus) is treated as insufficiently known and two (P. sanderianus, P. nitens) are treated as doubtful. Pothos latifolius L. is excluded from Araceae [= Piper sp.]. The following new synonymies are proposed: Pothos longipedunculatus Ridl. non Engl. = P. brevivaginatus; P. acuminatissimus = P. dolichophyllus; P. borneensis = P. insignis; P. scandens var. javanicus, P. macrophyllus and P. vrieseanus = P. junghuhnii; P. rumphii = P. tener; P. lorispathus = P. leptostachyus; P. kinabaluensis = P. longivaginatus; P. merrillii and P. ovatifolius var. simalurensis = P. ovatifolius; P. sumatranus, P. korthalsianus, P. inaequalis and P. jacobsonii = P. oxyphyllus. Relationships within Pothos and the taxonomic robustness of the satellite genera are discussed. Keys to the genera and species of Potheae and the subgenera and supergroups of Pothos for the region are provided. -

Laporan Tahunan Bank Maluku
2012 Laporan Tahunan Bank Maluku • BERUBAH MENJADI LEBIH BAIK • TRANSFORMED TO BE BETTER Bank Maluku telah memasuki era transformasi Bank Maluku has entered an era of transformation Era baru dalam pengembangan jati diri merubah a new era in the development of identity mimpi menjadi kenyataan change dream into reality Tak mudah merubah sebuah paradigma yang It is not easy to change a paradigm the common menjadi cita-cita bersama dengan dinamika yang goal with di erent dynamics berbeda Berubah Menjadi Lebih Baik Meniti asa menjadi Changed for the Better Climbing up into the Jawara di Maluku maupun Maluku Utara Sebagai champs in Maluku and North Maluku as Regional Champion Bank Regional Champion Bank. Laporan Tahunan PT Bank Pembangunan Daerah Maluku Tahun 2012 Annual Report 2012 PT Bank Pembangunan Daerah Maluku TRANSFORMED TO BE BETTER Daftar Isi Table of Content PEMBUKAAN 1 OPENING Nilai – Nilai Budaya Kerja PT. Bank Maluku 4 Value - Value of Cultural Work PT. Bank Maluku - Kepercayaan 4 - Confi dence - Integritas 4 - Integrity - Profesional 4 - Professional - Melayani 4 - Serve - Komitmen 4 - commitment - Disiplin 4 - Discipline - Kerjasama 4 - Cooperation Visi dan Misi 6-7 Vision and Mission Motto 8 Motto Makna Logo 9 Meaning Logo LAPORAN MANAJEMEN 10 MANAGEMENT REPORT Laporan Direksi 10 Report of the Board of Directors Laporan Dewan Komisaris 20 Report of the Board of Commissioners PROFIL PERUSAHAAN 26 COMPANY PROFILE Identitas Perusahaan 27 Company identity Sejarah Singkat 28 A Brief History Ikhtisar Keuangan 30 Financial Highlights -

The Transport Trend of Thailand and Malaysia
Executive Summary Report The Potential Assessment and Readiness of Transport Infrastructure and Services in Thailand for ASEAN Economic Community (AEC) Content Page 1. Introduction 1.1 Rationales 1 1.2 Objectives of Study 1 1.3 Scopes of Study 2 1.4 Methodology of Study 4 2. Current Status of Thailand Transport System in Line with Transport Agreement of ASEAN Community 2.1 Master Plan and Agreement on Transport System in ASEAN 5 2.2 Major Transport Systems for ASEAN Economic Community 7 2.2.1 ASEAN Highway Network 7 2.2.2 Major Railway Network for ASEAN Economic Community 9 2.2.3 Main Land Border Passes for ASEAN Economic Community 10 2.2.4 Main Ports for ASEAN Economic Community 11 2.2.5 Main Airports for ASEAN Economic Community 12 2.3 Efficiency of Current Transport System for ASEAN Economic Community 12 3. Performance of Thailand Economy and Transport Trend after the Beginning of ASEAN Economic Community 3.1 Factors Affecting Cross-Border Trade and Transit 14 3.2 Economic Development for Production Base Thriving in Thailand 15 3.2.1 The analysis of International Economic and Trade of Thailand and ASEAN 15 3.2.2 Major Production Bases and Commodity Flow of Prospect Products 16 3.2.3 Selection of Potential Industries to be the Common Production Bases of Thailand 17 and ASEAN 3.2.4 Current Situation of Targeted Industries 18 3.2.5 Linkage of Targeted Industries at Border Areas, Important Production Bases, 19 and Inner Domestic Areas TransConsult Co., Ltd. King Mongkut’s University of Technology Thonburi 2T Consulting and Management Co., Ltd. -

Transboundary Movements of Hazardous Wastes in THAILAND
Transboundary Movements of Hazardous Wastes IN THAILAND MS. PORNPIMON CHAREONSONG POLLUTION CONTROL DEPARTMENT Joint Meeting of Asian Network and REN November 20th, 2013, Bangkok, Thailand Background 2 Thailand has become a party of the Basel Convention since 1998. Competent Authority: Department of Industrial Works (DIW/MoI) Focal Point : Pollution Control Department (PCD/MNRE) Involved agencies : Customs Department/ Marine Department /Port Authority of Thailand/ Ministry of Foreign Affairs, etc. Coordinating mechanism : National Committee on Basel Convention Customs Houses Mae Sai Customs House Regional 1 = 11 Customs House Regional 2 = 9 Customs House Chiang Dao Customs House Regional 3 = 9 Customs House Chiang Saen Customs House Regional 4 = 18 Customs House Total = 47 Customs House 3 Chiang Khong Customs House Thung Chang Customs House Tha Li Customs House Mae Hong Son Customs House Chiang Mai Airport Customs House Nong Khai Customs House Chiang Khan Customs House Mae Sariang Customs House Bueng Kan Customs House Mae Sot Customs House Nakorn Phanom Customs House Mukdahan Customs House Khemarat Customs House Sangkhla Buri Customs House Chong Mek Customs House Chong Chom Customs House Aranyaprathet Customs House Mae Klong Customs House Maptaphut Customs House Prachuap Khiri Khan Customs House Chantaburi Customs House Chumporn Customs House Klong Yai Customs House Ranong Customs House Ban Don Customs House Ko Samui Customs House Phuket International Airport Customs House Phuket Customs House Sichon Customs House Nakhon Si Thammarat -

Bil. Nombor Nama Penternak / Lokasi Ladang. Alamat Surat Menyurat
Bil. Nombor Nama Penternak / Lokasi Ladang. Alamat Surat Menyurat. Nombor Tarikh Fi Ladang Kapasiti Catatan. Kod Ldg. Syarikat. Telefon. Luput. ( Rm ) ( Ekor ) 1 CA 069(D) Dato' Chow Liong. Lot 3751, Bintang, Mukim 3A Lorong SJ 1/3, Taman Emas, 09-4789309 RM500.00 42,000 ( Hiap Keong ) Luit, Maran. Kampong Sri Jaya, 28030 Maran. 2 CA 072(D) Dato' Chow Liong. PT 414, Ulu Sak, Jerantut 3A Lorong SJ 1/3, Taman Emas, 09-4789309 RM300.00 20,000 ( Hiap Keong ) Maran Road, Mukim Chenor, Kampong Sri Jaya, 28030 Maran. Maran. 3 CA 074(D) Dato' Chow Liong. Lot 1677, Sungai Kertam, 3A,Lorong SJ 1/3,Taman Emas, 09-4789309 RM500.00 42,000 ( Hiap Keong ) Mukim Luit, Maran. Kampong Sri Jaya, 28030 Maran. 4 CA 076(D) Dato' Chow Liong. Lot 8510, Kg. Sri Berjaya, 3A Lorong SJ 1/3, Taman Emas, 09-4789309 RM300.00 19,000 ( Hiap Keong ) Mukim Luit,Maran. Kampong Sri Jaya, 28030 Maran. 5 CA 077(D) Dato' Chow Liong. Lot 4194, 4199, PT 637, PT 3A Lorong SJ 1/3, Taman Emas, 09-4789309 RM500.00 120,000 ( Hiap Keong ) 638, Bukit Tunggal, Mukim Kampong Sri Jaya, 28030 Luit, Maran. Maran. 6 CA 079(D) Dato' Chow Liong. PT 1585, Sungai Kertam, 3A Lorong SJ 1/3, Taman Emas, 09-4789309 RM500.00 32,000 ( Hiap Keong ) Mukim Luit, Maran. Kampong Sri Jaya, 28030 Maran. 7 CA 249(D) Mohd Naim Bin Awang Lot 1208, Mukim Bukit No:7,Tingkat 2,Jalan Chengal 1/1, 09-2909773 RM500.00 33,000 (RISDA Smallholders Segumpal, Maran. -

This Thesis Has Been Submitted in Fulfilment of the Requirements for a Postgraduate Degree (E.G
This thesis has been submitted in fulfilment of the requirements for a postgraduate degree (e.g. PhD, MPhil, DClinPsychol) at the University of Edinburgh. Please note the following terms and conditions of use: This work is protected by copyright and other intellectual property rights, which are retained by the thesis author, unless otherwise stated. A copy can be downloaded for personal non-commercial research or study, without prior permission or charge. This thesis cannot be reproduced or quoted extensively from without first obtaining permission in writing from the author. The content must not be changed in any way or sold commercially in any format or medium without the formal permission of the author. When referring to this work, full bibliographic details including the author, title, awarding institution and date of the thesis must be given. Molecular Species Delimitation, Taxonomy and Biogeography of Sri Lankan Gesneriaceae Subhani Wathsala Ranasinghe Doctor of Philosophy The University of Edinburgh Royal Botanic Garden Edinburgh 2017 Declaration I hereby declare that the work contained in this thesis is my own unless otherwise acknowledged and cited. This thesis has not in whole or in part been previously presented for any degree Subhani Wathsala Ranasinghe 24th January 2017. i Abstract The plant family Gesneriaceae is represented in Sri Lanka by six genera: Aeschynanthus, Epithema, Championia, Henckelia, Rhynchoglossum and Rhynchotechum, with 13 species (plus one subspecies/variety) of which ten are endemic including the monotypic genus Championia, according to the last revision in 1981. They are exclusively distributed in undisturbed habitats, and some have high ornamental value. The species are morphologically diverse, but face a problem of taxonomic delineation, which is further complicated by the presence of putative hybrids. -
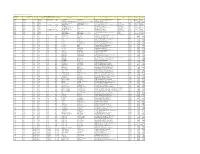
Colgate Palmolive List of Mills As of June 2018 (H1 2018) Direct
Colgate Palmolive List of Mills as of June 2018 (H1 2018) Direct Supplier Second Refiner First Refinery/Aggregator Information Load Port/ Refinery/Aggregator Address Province/ Direct Supplier Supplier Parent Company Refinery/Aggregator Name Mill Company Name Mill Name Country Latitude Longitude Location Location State AgroAmerica Agrocaribe Guatemala Agrocaribe S.A Extractora La Francia Guatemala Extractora Agroaceite Extractora Agroaceite Finca Pensilvania Aldea Los Encuentros, Coatepeque Quetzaltenango. Coatepeque Guatemala 14°33'19.1"N 92°00'20.3"W AgroAmerica Agrocaribe Guatemala Agrocaribe S.A Extractora del Atlantico Guatemala Extractora del Atlantico Extractora del Atlantico km276.5, carretera al Atlantico,Aldea Champona, Morales, izabal Izabal Guatemala 15°35'29.70"N 88°32'40.70"O AgroAmerica Agrocaribe Guatemala Agrocaribe S.A Extractora La Francia Guatemala Extractora La Francia Extractora La Francia km. 243, carretera al Atlantico,Aldea Buena Vista, Morales, izabal Izabal Guatemala 15°28'48.42"N 88°48'6.45" O Oleofinos Oleofinos Mexico Pasternak - - ASOCIACION AGROINDUSTRIAL DE PALMICULTORES DE SABA C.V.Asociacion (ASAPALSA) Agroindustrial de Palmicutores de Saba (ASAPALSA) ALDEA DE ORICA, SABA, COLON Colon HONDURAS 15.54505 -86.180154 Oleofinos Oleofinos Mexico Pasternak - - Cooperativa Agroindustrial de Productores de Palma AceiteraCoopeagropal R.L. (Coopeagropal El Robel R.L.) EL ROBLE, LAUREL, CORREDORES, PUNTARENAS, COSTA RICA Puntarenas Costa Rica 8.4358333 -82.94469444 Oleofinos Oleofinos Mexico Pasternak - - CORPORACIÓN