Zootaxa, Speleonectes Kakuki, a New Species of Remipedia (Crustacea) from Anchialine
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Coastal Circulation and Water-Column Properties in The
Coastal Circulation and Water-Column Properties in the War in the Pacific National Historical Park, Guam— Measurements and Modeling of Waves, Currents, Temperature, Salinity, and Turbidity, April–August 2012 Open-File Report 2014–1130 U.S. Department of the Interior U.S. Geological Survey FRONT COVER: Left: Photograph showing the impact of intentionally set wildfires on the land surface of War in the Pacific National Historical Park. Right: Underwater photograph of some of the healthy coral reefs in War in the Pacific National Historical Park. Coastal Circulation and Water-Column Properties in the War in the Pacific National Historical Park, Guam— Measurements and Modeling of Waves, Currents, Temperature, Salinity, and Turbidity, April–August 2012 By Curt D. Storlazzi, Olivia M. Cheriton, Jamie M.R. Lescinski, and Joshua B. Logan Open-File Report 2014–1130 U.S. Department of the Interior U.S. Geological Survey U.S. Department of the Interior SALLY JEWELL, Secretary U.S. Geological Survey Suzette M. Kimball, Acting Director U.S. Geological Survey, Reston, Virginia: 2014 For product and ordering information: World Wide Web: http://www.usgs.gov/pubprod Telephone: 1-888-ASK-USGS For more information on the USGS—the Federal source for science about the Earth, its natural and living resources, natural hazards, and the environment: World Wide Web: http://www.usgs.gov Telephone: 1-888-ASK-USGS Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government. Suggested citation: Storlazzi, C.D., Cheriton, O.M., Lescinski, J.M.R., and Logan, J.B., 2014, Coastal circulation and water-column properties in the War in the Pacific National Historical Park, Guam—Measurements and modeling of waves, currents, temperature, salinity, and turbidity, April–August 2012: U.S. -

Lozano-Fernandez Et Al
Citation for published version: Lozano-Fernandez, J, Giacomelli, M, Fleming, JF, Chen, A, Vinther, J, Thomsen, PF, Glenner, H, Palero, F, Legg, DA, Iliffe, TM, Pisani, D & Olesen, J 2019, 'Pancrustacean Evolution Illuminated by Taxon-Rich Genomic- Scale Data Sets with an Expanded Remipede Sampling', Genome biology and evolution, vol. 11, no. 8, pp. 2055-2070. https://doi.org/10.1093/gbe/evz097 DOI: 10.1093/gbe/evz097 Publication date: 2019 Link to publication University of Bath Alternative formats If you require this document in an alternative format, please contact: [email protected] General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. Take down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. Download date: 05. Oct. 2021 GBE Pancrustacean Evolution Illuminated by Taxon-Rich Genomic- Scale Data Sets with an Expanded Remipede Sampling 1,2,9,* 1 2,10 2,11 1,2 Jesus Lozano-Fernandez , Mattia Giacomelli , James F. Fleming ,AlbertChen , Jakob Vinther , Philip Downloaded from https://academic.oup.com/gbe/article-abstract/11/8/2055/5528088 by University of Cambridge user on 30 September 2019 Francis Thomsen3,12, Henrik Glenner4, Ferran Palero5,6,DavidA.Legg7,ThomasM.Iliffe8, Davide -

California's Coastal Sea Caves, NSS News, October 1998
California’s Coastal Sea Caves by Dave Bunnell Large chamber in Cave-in-Rock Cave, San Luis Obispo County elmets, lights, wetsuits, fins, face California, with its more accessible caves, are undoubtedly caves in Humboldt and Del masks and snorkels, vertical gear, has yielded as many again to the surveyor’s Norte counties, no survey work has been H and sea kayaks were not what I was tape and doubtless many more remain, undertaken there. I’ll begin with the southern used to packing for a cave trip until I especially in the remote northern counties. counties, for that was the original focus of moved to California. But after a couple of Much has been published over the years in sea cave survey. years of caving here, I realized that there were grotto and regional publications, but this is large numbers of caves here that had been the first time I’ve written an overall summary SAN DIEGO COUNTY largely ignored. After all, what caver wants of the Survey’s work on coastal caves. Perhaps the most famous of the sea caves to spend his weekends at the beach? on the California coast are the “seven caves” Systematic exploration and survey of GENERALITIES at La Jolla, formed in a 200’ high cliff of California’s many sea caves really began While not as large on average as the Cretaceous-age sandstone. A perennial when Carol Vesely and I chartered the island caves, the coastal caves are found in tourist favorite, La Jolla Cove and its cavey Southern California Sea Cave Survey in numerous host rocks that lend diverse cliffs are featured on dozens of old postcards. -

Biochemical Divergence Between Cavernicolous and Marine
The position of crustaceans within Arthropoda - Evidence from nine molecular loci and morphology GONZALO GIRIBET', STEFAN RICHTER2, GREGORY D. EDGECOMBE3 & WARD C. WHEELER4 Department of Organismic and Evolutionary- Biology, Museum of Comparative Zoology; Harvard University, Cambridge, Massachusetts, U.S.A. ' Friedrich-Schiller-UniversitdtJena, Instituifiir Spezielte Zoologie und Evolutionsbiologie, Jena, Germany 3Australian Museum, Sydney, NSW, Australia Division of Invertebrate Zoology, American Museum of Natural History, New York, U.S.A. ABSTRACT The monophyly of Crustacea, relationships of crustaceans to other arthropods, and internal phylogeny of Crustacea are appraised via parsimony analysis in a total evidence frame work. Data include sequences from three nuclear ribosomal genes, four nuclear coding genes, and two mitochondrial genes, together with 352 characters from external morphol ogy, internal anatomy, development, and mitochondrial gene order. Subjecting the com bined data set to 20 different parameter sets for variable gap and transversion costs, crusta ceans group with hexapods in Tetraconata across nearly all explored parameter space, and are members of a monophyletic Mandibulata across much of the parameter space. Crustacea is non-monophyletic at low indel costs, but monophyly is favored at higher indel costs, at which morphology exerts a greater influence. The most stable higher-level crusta cean groupings are Malacostraca, Branchiopoda, Branchiura + Pentastomida, and an ostracod-cirripede group. For combined data, the Thoracopoda and Maxillopoda concepts are unsupported, and Entomostraca is only retrieved under parameter sets of low congruence. Most of the current disagreement over deep divisions in Arthropoda (e.g., Mandibulata versus Paradoxopoda or Cormogonida versus Chelicerata) can be viewed as uncertainty regarding the position of the root in the arthropod cladogram rather than as fundamental topological disagreement as supported in earlier studies (e.g., Schizoramia versus Mandibulata or Atelocerata versus Tetraconata). -

Where America's Day Begins
Guam Where America’s Day Begins 1 Tumon Bay 2 Two Lovers Point 3 Local boy and mango 4 Where is Paradise? U.S. Territory located • RUSSIA in the Pacific Ocean, Moscow Vladivostok Beijing CHINA KOREA a true tropical island Seoul Osaka JAPAN KOREA Tokyo CHINA JAPAN Shanghai Commonwealth of the TAIWANNorthern Mariana Islands Situated approximately Hong Kong Taipei Saipan HONG KONG • Hanoi TAIWAN GUAM TERRITORY OF THE USA 900 miles north of the Manila PHILIPPINES PHILIPPINES Ho Chi Minh City equator VIETNAM • 32 miles long and ranges between 4 and 8 miles in width • Shaped like a footprint, Guam is the first step into Micronesia 5 Gateway to the Marianas and Micronesia The meaning of Micronesia: • COMMONWEALTH Saipan MICRONESIA OF NORTHERN Tinian MARIANA ISLANDS Rota “micro” – tiny Pacific Ocean Philippine Sea “nesia” – many GUAM TERRITORY OF U.S.A. REPUBLIC FEDERATED STATES REPUBLIC OF Micronesia – “many tiny OF PALAU OF MICRONESIA MARSHALL ISLANDS Pohnpei Kwajalein islands” Palau Yap Chuuk Kosrae Majuro • Largest and most populated island in the Marianas • Communications, transportation and financial hub in the Marianas 6 RUSSIA Getting Moscow Vladivostok Beijing CHINA KOREA Seoul Osaka JAPAN to Guam KOREA Tokyo CHINA JAPAN Shanghai Commonwealth of the TAIWANNorthern Mariana Islands From Hong Kong Taipei HONG KONG Hanoi TAIWAN GUAM TERRITORY OF THE USA Manila PHILIPPINES Vietnam PHILIPPINES Ho Chi Minh City VIETNAM Flight Time Hanoi to Guam via Korea 9 hrs via Taiwan 6 hrs 40 min via Tokyo 8 hrs 50 min Ho Chi Minh City via Hong Kong 10 hrs to Guam via Taiwan 7 hrs 15 min via Manila 6 hrs 30 min 7 Climate & Geography • Average 85ºF (29ºC) • Guam is on the edge of the Mariana Trench, the deepest trench in the world • Craggy limestone cliffs dominate in the North and rolling hills with spectacular views in the South 8 The People • 160,000 Residents • Population is 37% Chamorro, 26% Filipino and 11% other Pacific Islanders. -

Beaches Unit (4.5 Pts) Section
T. James Noyes, El Camino College Beaches Unit (Topic 6A) – page 1 Name: Beaches Unit (4.5 pts) Section: Beaches and Shorelines are always changing Waves are slowly and inexorably altering the shoreline, breaking down material in some places and carrying it to other places. Each wave has a small effect, but waves keep coming minute after minute, day after day, year after year. Significant changes in the shoreline (tens of feet) can occur within a human lifetime. For example, old maps of Encinitas (located along the coastal cliffs by Interstate 5 on the way down to San Diego) show that it has lost about a city block of land to the sea in the last century. In some places along the coast of Alaska, the shoreline has eroded 900 meters in 50 years, an average of about 18 meters (60 feet) per year! Beach Sediments Beach sediments are composed of whatever sediments are available locally: sand, cobbles, gravel, coral fragments, shell fragments, and so on. (In fact, tiny plastic sediments are making up a larger and larger component of our beaches.) Beach sediments are characterized by (1) the kind of material that they are made out of, (2) their size, (3) their shape, and (4) their sorting. (Well sorted sediments all have about the same size and shape, while poorly sorted sediments are a jumbled mix of sediments with many different sizes and shapes. Lithogenous sediments (“rock sediments”) are produced from the weathering of the rock of the land. Rocks are broken down into pieces (sediments) by the physical impact of water, wind, and other rocks; by chemicals dissolved in water; and repeated heating and cooling. -
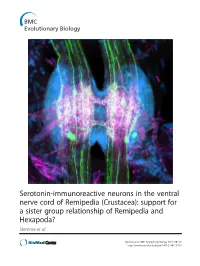
Serotonin-Immunoreactive Neurons in the Ventral Nerve Cord of Remipedia (Crustacea): Support for a Sister Group Relationship of Remipedia and Hexapoda? Stemme Et Al
Serotonin-immunoreactive neurons in the ventral nerve cord of Remipedia (Crustacea): support for a sister group relationship of Remipedia and Hexapoda? Stemme et al. Stemme et al. BMC Evolutionary Biology 2013, 13:119 http://www.biomedcentral.com/1471-2148/13/119 Stemme et al. BMC Evolutionary Biology 2013, 13:119 http://www.biomedcentral.com/1471-2148/13/119 RESEARCH ARTICLE Open Access Serotonin-immunoreactive neurons in the ventral nerve cord of Remipedia (Crustacea): support for a sister group relationship of Remipedia and Hexapoda? Torben Stemme1, Thomas M Iliffe2, Björn M von Reumont3, Stefan Koenemann4, Steffen Harzsch5 and Gerd Bicker1* Abstract Background: Remipedia were initially seen as a primitive taxon within Pancrustacea based on characters considered ancestral, such as the homonomously segmented trunk. Meanwhile, several morphological and molecular studies proposed a more derived position of Remipedia within Pancrustacea, including a sister group relationship to Hexapoda. Because of these conflicting hypotheses, fresh data are crucial to contribute new insights into euarthropod phylogeny. The architecture of individually identifiable serotonin-immunoreactive neurons has successfully been used for phylogenetic considerations in Euarthropoda. Here, we identified neurons in three species of Remipedia with an antiserum against serotonin and compared our findings to reconstructed ground patterns in other euarthropod taxa. Additionally, we traced neurite connectivity and neuropil outlines using antisera against acetylated α-tubulin and synapsin. Results: The ventral nerve cord of Remipedia displays a typical rope-ladder-like arrangement of separate metameric ganglia linked by paired longitudinally projecting connectives. The peripheral projections comprise an intersegmental nerve, consisting of two branches that fuse shortly after exiting the connectives, and the segmental anterior and posterior nerve. -

Adventure Sea Cave Kayak Tours on Santa Cruz Island at Scorpion Anchorage
ADVENTURE SEA CAVE KAYAK TOURS ON SANTA CRUZ ISLAND AT SCORPION ANCHORAGE THIS IS NOT A BOOKING CONFIRMATION. The information below is only for reference. Please make sure to reference your booking confirmation sent via email for exact times and locations. If you have any questions please contact the Channel Islands Adventure Company - we will be happy to help you. Paddling Time and Distance 2.5-3 miles, 2.5-3 hours of island kayaking Tour Level of Difficulty Beginner through advanced experience levels welcome. Children under 12 must ride in double kayak with a parent. Adventure Sea Cave Kayak Tour Itinerary Your morning will begin at the Island Packers ferry dock for check-in at either 7am or 8am, depending on which ferry departure time you select (8am departure or 9am departure). You’ll want to look for Channel Islands Adventure Company guides just outside of the Island Packers building. Your guide will have your tickets. Detailed driving directions will be sent upon booking in your confirmation email. The ferry ride is generally 1-1.5 hour(s) depending on wildlife sightings. You’ll then meet your guide(s) on the island at 10:30am.We provide a basic paddling instruction/safety talk and distribute all necessary gear before hitting the water around 11:15am. Afterward we begin the guided paddle along the coast in one direction, exploring the sea caves as you go. At some point you turn around and come back past the landing area and explore in the other direction. You’ll return to the beach around 2:00-2:30pm, and have until about 3:30–4:30pm (your ferry captain or guide will tell you your pick-up time) before leaving the island. -

NPWS Inishmore Island SAC (Site Code: 213) Conservation Objectives
NPWS Inishmore Island SAC (site code: 213) Conservation objectives supporting document - Marine Habitats Version 1 January 2015 Introduction Inishmore Island SAC is designated for the marine Annex I qualifying interests of Reefs and Submerged or partially submerged sea caves (Figures 1 and 2). A BioMar survey of this site was carried out in 1993 (Picton and Costello, 1997), a seacave survey and intertidal and subtidal reef surveys were undertaken in 2012 (MERC, 2012 and MERC, 2013); these data were used to determine the physical and biological nature of this SAC. The distribution and ecology of intertidal or subtidal seacaves has not previously been the subject of scientific investigation in Ireland and the extents of very few individual caves have been mapped in detail. Whilst surveys undertaken in the UK indicate the structure and functions of seacaves are largely influenced by hydrodynamic forces and water quality, no such information is yet available for Ireland. Aspects of the biology and ecology of the Annex I habitat are provided in Section 1. The corresponding site-specific conservation objectives will facilitate Ireland delivering on its surveillance and reporting obligations under the EU Habitats Directive (92/43/EC). Ireland also has an obligation to ensure that consent decisions concerning operations/activities planned for Natura 2000 sites are informed by an appropriate assessment where the likelihood of such operations or activities having a significant effect on the SAC cannot be excluded. Further ancillary information concerning the practical application of the site-specific objectives and targets in the completion of such assessments is provided in Section 2. -

A Silurian Soft-Bodied Biota Author(S): Donald G
A Silurian Soft-Bodied Biota Author(s): Donald G. Mikulic, Derek E. G. Briggs, Joanne Kluessendorf Source: Science, New Series, Vol. 228, No. 4700 (May 10, 1985), pp. 715-717 Published by: American Association for the Advancement of Science Stable URL: http://www.jstor.org/stable/1694543 Accessed: 24/02/2010 21:52 Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at http://www.jstor.org/action/showPublisher?publisherCode=aaas. Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. American Association for the Advancement of Science is collaborating with JSTOR to digitize, preserve and extend access to Science. http://www.jstor.org cept where infilled by diagenetic fluor- apatite. -

Diving in Guam
Diving in Guam Introduction Popular Outer Reef Sites Guam’s tropical waters are teaming with hundreds Guam is home to one of the world’s most amazing of varieties of colorful fish and coral species, a seascape of underwater phenomena, the Blue Hole, a natural 300-foot unimaginable beauty and clarity, with historic wrecks from shaft cut into the reef off Orote Point. A plethora of marine World War I and World War II. life and coral also inhabit several other outer reef dive sites including the Crevice, Coral Gardens, Shark Pit, Barracuda Guam’s open ocean is not only home to diverse coral and Rock, Hap’s Reef, and the elusive 11-Mile Reef. marine life; it’s one of Micronesia’s most affordable places to dive. Home to a variety of dives for all certification levels, the • Blue Hole island’s waters stay from 82-86°F year-round, with visibility One of Guam’s most unique dive sites, the Blue Hole is up to 150 feet. Dive environments for beginners and more located toward the tip of Orote Peninsula on the southern experienced divers are concentrated more heavily in the coast. Characterized as a 300-foot shaft cut into reef made south, with the exception of a few accessible dive spots in of limestone, the site offers excellent visibility that often northern Guam. allows divers to see the top of the hole from the surface. The shaft actually drops much further than air diving allows, but a large window opens to the outer wall at about 125 feet, allowing divers to exit and ascend after a free fall through the shaft. -

Fuxianhuiid Ventral Nerve Cord and Early Nervous System Evolution in Panarthropoda
Fuxianhuiid ventral nerve cord and early nervous system evolution in Panarthropoda Jie Yanga, Javier Ortega-Hernándezb,1, Nicholas J. Butterfieldb, Yu Liua,c,d, George S. Boyanc, Jin-bo Houa, Tian Lane, and Xi-guang Zhanga,2 aYunnan Key Laboratory for Paleobiology, Yunnan University, Kunming 650091, China; bDepartment of Earth Sciences, University of Cambridge, Cambridge CB2 3EQ, United Kingdom; cDevelopmental Neurobiology Group, Biocenter, Ludwig-Maximilians-Universität, 82152 Martinsried, Germany; dGeoBio-Center Ludwig-Maximilians-Universität, Munich 80333, Germany; and eCollege of Resources and Environmental Engineering, Guizhou University, Guiyang 550003, China Edited by Gregory D. Edgecombe, The Natural History Museum, London, United Kingdom, and accepted by the Editorial Board January 29, 2016 (received for review November 14, 2015) Panarthropods are typified by disparate grades of neurological feature as the VNC. This interpretation is supported by com- organization reflecting a complex evolutionary history. The fossil parisons with other preserved components of the internal anat- record offers a unique opportunity to reconstruct early character omy. For instance, the VNC can be readily distinguished from evolution of the nervous system via exceptional preservation in the digestive tract of C. kunmingensis, which is expressed as a extinct representatives. Here we describe the neurological archi- comparatively larger (maximum width, ∼860 μm) but fully tecture of the ventral nerve cord (VNC) in the upper-stem group compressed, linear structure running almost the entire length of euarthropod Chengjiangocaris kunmingensis from the early Cam- the animal (23, figure 1 d and e). The VNC extends from at least brian Xiaoshiba Lagerstätte (South China). The VNC of C. kunmin- the five anteriormost reduced trunk tergites (i.e., dorsal exo- gensis comprises a homonymous series of condensed ganglia that skeletal plates) to tergite T23 at the posterior end of the trunk extend throughout the body, each associated with a pair of bi- (Figs.