Characterization of the Novel Defective Cyp2c9*24
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Identification and Developmental Expression of the Full Complement Of
Goldstone et al. BMC Genomics 2010, 11:643 http://www.biomedcentral.com/1471-2164/11/643 RESEARCH ARTICLE Open Access Identification and developmental expression of the full complement of Cytochrome P450 genes in Zebrafish Jared V Goldstone1, Andrew G McArthur2, Akira Kubota1, Juliano Zanette1,3, Thiago Parente1,4, Maria E Jönsson1,5, David R Nelson6, John J Stegeman1* Abstract Background: Increasing use of zebrafish in drug discovery and mechanistic toxicology demands knowledge of cytochrome P450 (CYP) gene regulation and function. CYP enzymes catalyze oxidative transformation leading to activation or inactivation of many endogenous and exogenous chemicals, with consequences for normal physiology and disease processes. Many CYPs potentially have roles in developmental specification, and many chemicals that cause developmental abnormalities are substrates for CYPs. Here we identify and annotate the full suite of CYP genes in zebrafish, compare these to the human CYP gene complement, and determine the expression of CYP genes during normal development. Results: Zebrafish have a total of 94 CYP genes, distributed among 18 gene families found also in mammals. There are 32 genes in CYP families 5 to 51, most of which are direct orthologs of human CYPs that are involved in endogenous functions including synthesis or inactivation of regulatory molecules. The high degree of sequence similarity suggests conservation of enzyme activities for these CYPs, confirmed in reports for some steroidogenic enzymes (e.g. CYP19, aromatase; CYP11A, P450scc; CYP17, steroid 17a-hydroxylase), and the CYP26 retinoic acid hydroxylases. Complexity is much greater in gene families 1, 2, and 3, which include CYPs prominent in metabolism of drugs and pollutants, as well as of endogenous substrates. -

Synonymous Single Nucleotide Polymorphisms in Human Cytochrome
DMD Fast Forward. Published on February 9, 2009 as doi:10.1124/dmd.108.026047 DMD #26047 TITLE PAGE: A BIOINFORMATICS APPROACH FOR THE PHENOTYPE PREDICTION OF NON- SYNONYMOUS SINGLE NUCLEOTIDE POLYMORPHISMS IN HUMAN CYTOCHROME P450S LIN-LIN WANG, YONG LI, SHU-FENG ZHOU Department of Nutrition and Food Hygiene, School of Public Health, Peking University, Beijing 100191, P. R. China (LL Wang & Y Li) Discipline of Chinese Medicine, School of Health Sciences, RMIT University, Bundoora, Victoria 3083, Australia (LL Wang & SF Zhou). 1 Copyright 2009 by the American Society for Pharmacology and Experimental Therapeutics. DMD #26047 RUNNING TITLE PAGE: a) Running title: Prediction of phenotype of human CYPs. b) Author for correspondence: A/Prof. Shu-Feng Zhou, MD, PhD Discipline of Chinese Medicine, School of Health Sciences, RMIT University, WHO Collaborating Center for Traditional Medicine, Bundoora, Victoria 3083, Australia. Tel: + 61 3 9925 7794; fax: +61 3 9925 7178. Email: [email protected] c) Number of text pages: 21 Number of tables: 10 Number of figures: 2 Number of references: 40 Number of words in Abstract: 249 Number of words in Introduction: 749 Number of words in Discussion: 1459 d) Non-standard abbreviations: CYP, cytochrome P450; nsSNP, non-synonymous single nucleotide polymorphism. 2 DMD #26047 ABSTRACT Non-synonymous single nucleotide polymorphisms (nsSNPs) in coding regions that can lead to amino acid changes may cause alteration of protein function and account for susceptivity to disease. Identification of deleterious nsSNPs from tolerant nsSNPs is important for characterizing the genetic basis of human disease, assessing individual susceptibility to disease, understanding the pathogenesis of disease, identifying molecular targets for drug treatment and conducting individualized pharmacotherapy. -

Supplementary Material
Supplementary Material Table S1: Significant downregulated KEGGs pathways identified by DAVID following exposure to five cinnamon- based phenylpropanoids (p < 0.05). p-value Term: Genes (Benjamini) Cytokine-cytokine receptor interaction: FASLG, TNFSF14, CXCL11, IL11, FLT3LG, CCL3L1, CCL3L3, CXCR6, XCR1, 2.43 × 105 RTEL1, CSF2RA, TNFRSF17, TNFRSF14, CCNL2, VEGFB, AMH, TNFRSF10B, INHBE, IFNB1, CCR3, VEGFA, CCR2, IL12A, CCL1, CCL3, CXCL5, TNFRSF25, CCR1, CSF1, CX3CL1, CCL7, CCL24, TNFRSF1B, IL12RB1, CCL21, FIGF, EPO, IL4, IL18R1, FLT1, TGFBR1, EDA2R, HGF, TNFSF8, KDR, LEP, GH2, CCL13, EPOR, XCL1, IFNA16, XCL2 Neuroactive ligand-receptor interaction: OPRM1, THRA, GRIK1, DRD2, GRIK2, TACR2, TACR1, GABRB1, LPAR4, 9.68 × 105 GRIK5, FPR1, PRSS1, GNRHR, FPR2, EDNRA, AGTR2, LTB4R, PRSS2, CNR1, S1PR4, CALCRL, TAAR5, GABRE, PTGER1, GABRG3, C5AR1, PTGER3, PTGER4, GABRA6, GABRA5, GRM1, PLG, LEP, CRHR1, GH2, GRM3, SSTR2, Chlorogenic acid Chlorogenic CHRM3, GRIA1, MC2R, P2RX2, TBXA2R, GHSR, HTR2C, TSHR, LHB, GLP1R, OPRD1 Hematopoietic cell lineage: IL4, CR1, CD8B, CSF1, FCER2, GYPA, ITGA2, IL11, GP9, FLT3LG, CD38, CD19, DNTT, 9.29 × 104 GP1BB, CD22, EPOR, CSF2RA, CD14, THPO, EPO, HLA-DRA, ITGA2B Cytokine-cytokine receptor interaction: IL6ST, IL21R, IL19, TNFSF15, CXCR3, IL15, CXCL11, TGFB1, IL11, FLT3LG, CXCL10, CCR10, XCR1, RTEL1, CSF2RA, IL21, CCNL2, VEGFB, CCR8, AMH, TNFRSF10C, IFNB1, PDGFRA, EDA, CXCL5, TNFRSF25, CSF1, IFNW1, CNTFR, CX3CL1, CCL5, TNFRSF4, CCL4, CCL27, CCL24, CCL25, CCL23, IFNA6, IFNA5, FIGF, EPO, AMHR2, IL2RA, FLT4, TGFBR2, EDA2R, -

Clinical Implications of 20-Hydroxyeicosatetraenoic Acid in the Kidney, Liver, Lung and Brain
1 Review 2 Clinical Implications of 20-Hydroxyeicosatetraenoic 3 Acid in the Kidney, Liver, Lung and Brain: An 4 Emerging Therapeutic Target 5 Osama H. Elshenawy 1, Sherif M. Shoieb 1, Anwar Mohamed 1,2 and Ayman O.S. El-Kadi 1,* 6 1 Faculty of Pharmacy and Pharmaceutical Sciences, University of Alberta, Edmonton T6G 2E1, AB, Canada; 7 [email protected] (O.H.E.); [email protected] (S.M.S.); [email protected] (A.M.) 8 2 Department of Basic Medical Sciences, College of Medicine, Mohammed Bin Rashid University of 9 Medicine and Health Sciences, Dubai, United Arab Emirates 10 * Correspondence: [email protected]; Tel.: 780-492-3071; Fax: 780-492-1217 11 Academic Editor: Kishor M. Wasan 12 Received: 12 January 2017; Accepted: 15 February 2017; Published: 20 February 2017 13 Abstract: Cytochrome P450-mediated metabolism of arachidonic acid (AA) is an important 14 pathway for the formation of eicosanoids. The ω-hydroxylation of AA generates significant levels 15 of 20-hydroxyeicosatetraenoic acid (20-HETE) in various tissues. In the current review, we discussed 16 the role of 20-HETE in the kidney, liver, lung, and brain during physiological and 17 pathophysiological states. Moreover, we discussed the role of 20-HETE in tumor formation, 18 metabolic syndrome and diabetes. In the kidney, 20-HETE is involved in modulation of 19 preglomerular vascular tone and tubular ion transport. Furthermore, 20-HETE is involved in renal 20 ischemia/reperfusion (I/R) injury and polycystic kidney diseases. The role of 20-HETE in the liver is 21 not clearly understood although it represents 50%–75% of liver CYP-dependent AA metabolism, 22 and it is associated with liver cirrhotic ascites. -

Rare Novel CYP2U1 and ZFYVE26 Variants Identified in Two Pakistani Families with Spastic 2 Paraplegia
1 Rare novel CYP2U1 and ZFYVE26 variants identified in two Pakistani families with spastic 2 paraplegia. 3 Farah Bibi1, Stephanie Efthymiou2, Thomas Bourinaris2, Ambreen Tariq2, SYNaPS Study Group, 4 Faisal Zafar3, Nouzhat Rana3, Vincenzo Salpietro2, Henry Houlden2, Ghazala Kaukab Raja1 5 6 1. University Institute of Biochemistry & Biotechnology, PMAS – Arid Agriculture University, 7 Rawalpindi, 43600, Pakistan 8 2. Department of Neuromuscular disorders, UCL Institute of Neurology, Queen Square, London 9 WC1N 3BG, UK 10 3. Department of Pediatrics, Multan Nishtar Hospital, Multan, 60000, Pakistan. 11 Corresponding author: Prof. Ghazala Kaukab Raja, University Institute of Biochemistry & 12 Biotechnology, PMAS-Arid Agriculture University, Rawalpindi, 43600, Pakistan, 13 [email protected] 14 Keywords: spastic paraplegia, SPG15, SPG56, next generation sequencing, consanguinity 15 Word count: 1894 16 17 Abstract 18 Bakground: Hereditary Spastic paraplegias (HSPs) are a clinically and genetically heterogeneous 19 group of degenerative disorders characterized by progressive spasticity and weakness of the 20 lower limbs. This study aimed to identify causative gene variants in two unrelated 21 consanguineous Pakistani families presented with 2 different forms of HSP. 22 Methods: Whole exome sequencing (WES) was performed in the two families and variants were 23 validated by Sanger sequencing and segregation analysis. 24 Analysis: In family A, a homozygous pathogenic variant in ZFYVE26 was identified in one family. 25 While in family B, a frameshift variant in CYP2U1 was identified in 4 affected individuals 26 presented with clinical features of SPG56. Our study is the first report of ZFYVE26 mutations 27 causing HSP in the Pakistani population and the second report of CYP2U1 in a Pakistani family. -

Metabolic Activation and Toxicological Evaluation of Polychlorinated Biphenyls in Drosophila Melanogaster T
www.nature.com/scientificreports OPEN Metabolic activation and toxicological evaluation of polychlorinated biphenyls in Drosophila melanogaster T. Idda1,7, C. Bonas1,7, J. Hofmann1, J. Bertram1, N. Quinete1,2, T. Schettgen1, K. Fietkau3, A. Esser1, M. B. Stope4, M. M. Leijs3, J. M. Baron3, T. Kraus1, A. Voigt5,6 & P. Ziegler1* Degradation of polychlorinated biphenyls (PCBs) is initiated by cytochrome P450 (CYP) enzymes and includes PCB oxidation to OH-metabolites, which often display a higher toxicity than their parental compounds. In search of an animal model refecting PCB metabolism and toxicity, we tested Drosophila melanogaster, a well-known model system for genetics and human disease. Feeding Drosophila with lower chlorinated (LC) PCB congeners 28, 52 or 101 resulted in the detection of a human-like pattern of respective OH-metabolites in fy lysates. Feeding fies high PCB 28 concentrations caused lethality. Thus we silenced selected CYPs via RNA interference and analyzed the efect on PCB 28-derived metabolite formation by assaying 3-OH-2′,4,4′-trichlorobiphenyl (3-OHCB 28) and 3′-OH-4′,4,6′-trichlorobiphenyl (3′-OHCB 28) in fy lysates. We identifed several drosophila CYPs (dCYPs) whose knockdown reduced PCB 28-derived OH-metabolites and suppressed PCB 28 induced lethality including dCYP1A2. Following in vitro analysis using a liver-like CYP-cocktail, containing human orthologues of dCYP1A2, we confrm human CYP1A2 as a PCB 28 metabolizing enzyme. PCB 28-induced mortality in fies was accompanied by locomotor impairment, a common phenotype of neurodegenerative disorders. Along this line, we show PCB 28-initiated caspase activation in diferentiated fy neurons. -

Original Article Cytochrome P450 Family Proteins As Potential Biomarkers for Ovarian Granulosa Cell Damage in Mice with Premature Ovarian Failure
Int J Clin Exp Pathol 2018;11(8):4236-4246 www.ijcep.com /ISSN:1936-2625/IJCEP0080020 Original Article Cytochrome P450 family proteins as potential biomarkers for ovarian granulosa cell damage in mice with premature ovarian failure Jiajia Lin1, Jiajia Zheng1, Hu Zhang1, Jiulin Chen1, Zhihua Yu1, Chuan Chen1, Ying Xiong3, Te Liu1,2 1Shanghai Geriatric Institute of Chinese Medicine, Longhua Hospital, Shanghai University of Traditional Chinese Medicine, Shanghai, China; 2Department of Pathology, Yale UniversitySchool of Medicine, New Haven, USA; 3Department of Gynaecology and Obestetrics, Xinhua Hospital Affiliated to Shanghai Jiaotong University School of Medicine, Shanghai, China Received May 21, 2018; Accepted June 29, 2018; Epub August 1, 2018; Published August 15, 2018 Abstract: Premature ovarian failure (POF) is the pathological aging of ovarian tissue. We have previously established a cyclophosphamide-induced mouse POF model and found that cyclophosphamide caused significant damage and apoptosis of mouse ovarian granulosa cells (mOGCs). To systematically explore the molecular biologic evidence of cyclophosphamide-induced mOGC damage at the gene transcription level, RNA-Seqwas used to analyse the differ- ences in mOGC transcriptomes between POF and control (PBS) mice. The sequencing results showed that there were 18765 differential transcription genes between the two groups, of which 192 were significantly up-regulated (log2 [POF/PBS] > 2.0) and 116 were significantly down-regulated (log2 [POF/PBS] < -4.0). Kyoto Encyclopedia of Genes and Genomes analysis found that the neuroactive ligand-receptor interaction pathway was significantly up-regulated and metabolic pathways were significantly down-regulated in the POF group. Gene Ontology analy- sis showed that the expression of plasma membrane, regulation of transcription and ion binding functions were significantly up-regulated in the POF group, while the expression of cell and cell parts, catalytic activity and single- organism process functions were significantly down-regulated. -

Screening 20-Hete Inhibitors in Microsomal Incubates
SCREENING 20-HETE INHIBITORS IN MICROSOMAL INCUBATES USING UPLC-MS/MS by Chenxiao Tang Bachelor of Science, Capital Medical University, 2014 Submitted to the Graduate Faculty of School of Pharmacy in partial fulfillment of the requirements for the degree of Master of Science University of Pittsburgh 2016 Copyright © by Chenxiao Tang 2016 ii SCREENING 20-HETE INHIBITORS IN MICROSOMAL INCUBATES USING UPLC-MS/MS Chenxiao Tang, B.S. University of Pittsburgh, 2016 20-hydroxyeicosatetraenoic acid (20-HETE) is a metabolite of arachidonic acid (AA) formed by cytochrome P450 (CYP) 4A11 and CYP4F2 in humans, with potent microvascular constriction activity. Inhibition of 20-HETE formation is neuroprotective in subarachnoid hemorrhage, cardiac arrest and thromboembolic stroke preclinical models. These findings suggest that inhibition of 20-HETE formation is a potential therapeutic strategy for neuroprotection after brain injury. At this point, a clinically relevant 20-HETE inhibitor is not available to be evaluated as a therapeutic intervention. Our goal is to identify a selective, metabolically stable, and potent 20-HETE inhibitor. Test compounds were obtained either via virtual screening against a CYP4F2 homology model or from scaffold hopping from structures of known inhibitors. Four different types of microsomes including human liver microsome (HLM), recombinant CYP4F2 (rCYP4F2), rat liver microsome (RLM) and rat kidney microsome (RKM) were used. AA was incubated in microsomes with/without compound for 20 min. 20-HETE formation rate was quantified using a validated UPLC-MS/MS assay and normalized by vehicle control group. Other eicosanoids including 15-, 12-HETEs, epoxyeicosatrienoic acids (EETs), iii and dihydroxyeicosatrienoic acids (DiHETs) were monitored simultaneously. -

Expression of CYP2S1 in Human Hepatic Stellate Cells
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Elsevier - Publisher Connector FEBS Letters 581 (2007) 781–786 Expression of CYP2S1 in human hepatic stellate cells Carylyn J. Mareka, Steven J. Tuckera, Matthew Korutha, Karen Wallacea,b, Matthew C. Wrighta,b,* a School of Medical Sciences, Institute of Medical Science, University of Aberdeen, Aberdeen, UK b Liver Faculty Research Group, School of Clinical and Laboratory Sciences, University of Newcastle, Newcastle, UK Received 22 November 2006; revised 16 January 2007; accepted 23 January 2007 Available online 2 February 2007 Edited by Laszlo Nagy the expression and accumulation of scarring extracellular Abstract Activated stellate cells are myofibroblast-like cells associated with the generation of fibrotic scaring in chronically fibrotic matrix protein [2]. It is currently thought an inhibition damaged liver. Gene chip analysis was performed on cultured fi- of fibrosis in liver diseases may be an effective approach to brotic stellate cells. Of the 51 human CYP genes known, 13 treating patients for which the cause is refractive to current CYP and 5 CYP reduction-related genes were detected with 4 treatments (e.g. in approx. 30% of hepatitis C infected CYPs (CYP1A1, CYP2E1, CY2S1 and CYP4F3) consistently patients) [2,3]. At present, there is no approved treatment for present in stellate cells isolated from three individuals. Quantita- fibrosis. tive RT-PCR indicated that CYP2S1 was a major expressed Inadvertent toxicity of drugs is often associated with a ‘‘met- CYP mRNA transcript. The presence of a CYP2A-related pro- abolic activation’’ by CYPs [1]. -
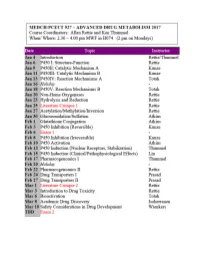
CYTOCHROME P450: Structure-Function
MEDCH 527 AER Jan. 4-6, 2017 CYTOCHROME P450: Structure-Function 1. General P450 Characteristics anD Taxonomy 2. Human P450s – Substrate anD Inhibitor Selectivities 3. Structure-Function Aspects of LiganD BinDing, P450 ReDuction anD Oxygen Activation References P450 Homepage -http:// http://drnelson.uthsc.edu/CytochromeP450.html Nelson DR et al., Pharmacogenetics. 1996 6(1):1-42: Nelson DR et al., Pharmacogenetics. 2004 14(1):1-18. Testa, B. The Biochemistry of Drug Metabolism: A 6 Part Series in Chem. BioDivers. (2006-2008). Sligar, SG. Glimpsing the critical intermediate in cytochrome P450 oxidations. Science. 2010 Nov 12;330(6006):924-5. Johnson EF, et al. Correlating structure and function of drug metabolizing enzymes:Progress and ongoing challenges. Drug Metab. Dispos. 42:9-22 (2014). Zientek MA and Youdim K, Reaction Phenotyping:Advances in experimental strategies used to characterize the contribution of drug metabolizing enzymes. Drug Metab. Dispos. 43:163-181 (2015). Almazroo et al., Drug Metabolism in the Liver. Clin. Liver Dis. 21:1-20 (2017). Absorption, Distribution, Metabolism and Excretion (ADME) of Orally Administered Drugs To reach their sites of action in the body, orally administered drugs must be absorbed from the small intestine, survive first pass metabolism - typically in the liver - before eventually being excreted, usually as drug metabolites in the bile and kidney. Therefore, clinically useful drugs must be able to cross an array of cell membranes, which are composed of a lipid bilayer. Drugs must exhibit an adequate degree of lipophilicity (logP of ~2-4) in order to able to dissolve into this lipoidal environment. Many drug metabolism processes render lipophilic drugs more water-soluble so as to facilitate excretion via the kidneys and bile. -

Biodiversity of P-450 Monooxygenase: Cross-Talk
Cytochrome P450: Oxygen activation and biodiversty 1 Biodiversity of P-450 monooxygenase: Cross-talk between chemistry and biology Heme Fe(II)-CO complex 450 nm, different from those of hemoglobin and other heme proteins 410-420 nm. Cytochrome Pigment of 450 nm Cytochrome P450 CYP3A4…. 2 High Energy: Ultraviolet (UV) Low Energy: Infrared (IR) Soret band 420 nm or g-band Mb Fe(II) ---------- Mb Fe(II) + CO - - - - - - - Visible region Visible bands Q bands a-band, b-band b a 3 H2O/OH- O2 CO Fe(III) Fe(II) Fe(II) Fe(II) Soret band at 420 nm His His His His metHb deoxy Hb Oxy Hb Carbon monoxy Hb metMb deoxy Mb Oxy Mb Carbon monoxy Mb H2O/Substrate O2-Substrate CO Substrate Soret band at 450 nm Fe(III) Fe(II) Fe(II) Fe(II) Cytochrome P450 Cys Cys Cys Cys Active form 4 Monooxygenase Reactions by Cytochromes P450 (CYP) + + RH + O2 + NADPH + H → ROH + H2O + NADP RH: Hydrophobic (lipophilic) compounds, organic compounds, insoluble in water ROH: Less hydrophobic and slightly soluble in water. Drug metabolism in liver ROH + GST → R-GS GST: glutathione S-transferase ROH + UGT → R-UG UGT: glucuronosyltransferaseGlucuronic acid Insoluble compounds are converted into highly hydrophilic (water soluble) compounds. 5 Drug metabolism at liver: Sleeping pill, pain killer (Narcotic), carcinogen etc. Synthesis of steroid hormones (steroidgenesis) at adrenal cortex, brain, kidney, intestine, lung, Animal (Mammalian, Fish, Bird, Insect), Plants, Fungi, Bacteria 6 NSAID: non-steroid anti-inflammatory drug 7 8 9 10 11 Cytochrome P450: Cysteine-S binding to Fe(II) heme is important for activation of O2. -

Arachidonic Acid Metabolism Zebrafish Mouse
Arachidonic Acid Metabolism Zebrafish Eicosanoids A. Nodes Edges 5,6-EET ephx2 5,6-DHET node 8 hpl 16hpl compound cyp2p10 8,9-EET ephx2 8,9-DHET node complex 14,15-EET ephx2 C14775 enrichment score -1 0 1 Linoleic acid metaolism 20-OH-Leukotriene Arachidonic 12(S)-HPETE 5(S)-HETE Leukotriene B4 cyp4f3 B4 acid alox12 gpx1a lta4h pla2g6 Leukotriene alox5a 5-HPETE alox5a A4 ltc4s Leukotriene C4 ggt5a Leukotriene D4 ptgs2a Prostaglandin Phosphatidylcholine G2 Prostaglandin I2 ptgis ptgs2a K17687 Prostaglandin E2 Prostaglandin ptges cyp4f3 20-HETE H2 Thromboxane cbr1 A2 cyp2u1 tbxas1 Prostaglandin F2 cbr1 alpha ptgdsb Prostaglandin D2 K07415 fam213b K17728 K17721 19(S)-HETE K17687 cyp2p10 cyp2u1 K07425 11,12-EET ephx2 11,12-DHET cyp2p10 Mouse B. Eicosanoids 5,6-EET Ephx2 5,6-DHET Nodes Edges rd1/rd1 -/- compound 8,9-EET Ephx2 8,9-DHET node Pde6b Rho Cyp2j6 node 14,15-EET Ephx2 C14775 complex Cyp2c29 16(R)-HETE enrichment score -1 0 1 Alox8 8(S)-HPETE Alox12b 12(R)-HPETE C14781 Arachidonic Cyp2j6 Cyp2j6 C14782 acid Linoleic acid Pla2g5 metaolism 12(S)-HPETE Cyp2j6 C14813 Cyp2j6 C14814 Alox12 15(S)-HPETE Gpx1 15(S)-HETE Phosphatidylcholine Alox15 20-OH-Leukotriene Alox5 5-HPETE Gpx1 5(S)-HETE Leukotriene B4 Cyp4f14 B4 Lta4h Ptgs1 Prostaglandin Leukotriene G2 Alox5 A4 Ltc4s Leukotriene C4 Ggt1 Leukotriene D4 Cyp4a10 Prostaglandin I2 Ptgis Cyp4f14 20-HETE Ptgs1 Prostaglandin E2 Cyp2u1 Prostaglandin Ptges H2 Thromboxane Cbr1 Tbxas1 A2 Cyp2e1 Cbr1 Prostaglandin F2 alpha K17728 Ptgdsb Prostaglandin D2 K17721 Fam213b 19(S)-HETE Cyp4a10 Cyp2j6 Cyp2u1 Cyp4a10 11,12-EET Ephx2 11,12-DHET Cyp2j6 Downloaded from iovs.arvojournals.org on 10/03/2021 Figure S1.