Sexual Dimorphism in Jurassic Ammonites
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Felix Gradstein, Lames Ogg and Alan Smith 18 the Jurassic Period J
Felix Gradstein, lames ogg and Alan smith 18 The Jurassic Period J. G. OGG iographic distribution of Jurassic GSSPs that have been ratified (ye Table 18.1 for more extensive listing). GSSPs for the honds) or are candidates (squares) on a mid-Jurassic map base-Jurassic, Late Jurassic stages, and some Middle Jurassic stages PNS in January 2004; see Table 2.3). Overlaps in Europe have are undefined. The projection center is at 30" E to place the center kured some GSSPs, and not all candidate sections are indicated of the continents in the center of the map. basaurs dominated theland surface. Ammonites are themain fossils neously considered his unit to he older. Alexander Brongniart rmrrelatingmarine deposits. Pangea supercontinent began to break (1829) coined the term "Terrains Jurassiques" when correlat- h md at the end of the Middle Jurassic the Central Atlantic was ing the "Jura Kalkstein" to the Lower Oolite Series (now as- m. Organic-rich sediments in several locations eventually became signed to Middle Jurassic) of the British succession. Leopold t source rocks helping to fuel modern civilization. von Buch (1839) established a three-fold subdivision for the Jurassic. The basic framework of von Buch has been retained as the three Jurassic series, although the nomenclature has var- 8.1 HISTORY AND SUBDIVISIONS ied (Black-Brown-White, Lias-Dogger-Malm, and currently L1.1 Overview of the Jurassic Lower-Middle-Upper). The immense wealth of fossils, particularly ammonites, in hc term "Jura Kalkstein" was applied by Alexander von the Jurassic strata of Britain, France, German5 and Switzer- bmholdt (1799) to a series ofcarhonate shelfdeposits exposed land was a magnet for innovative geologists, and modern con- the mountainous Jura region of northernmost Switzerland, cepts of hiostratigraphy, chronostratigraphy, correlation, and d he first recognized that these strata were distinct from paleogeography grew out of their studies. -

Back Matter (PDF)
GENERAL INDEX TO THE QUARTERLY JOURNAL AND PROCEEDINGS OF THE GEOLOGIC&L SOCIETY. AC~.AND,. H. D., on Vole. Series nr. Agglomerate, vole., of Lambay I., Herefordshire Beacon, 556-562 w. 141. maps & sect. Airolo (St: Gothard), Tremola Schists Aclisina, observats, on genus, 45"72 nr., 358, 359. & pls. iii-v. Aldeburgh (Suffolk), struct, of Cor. aciculata, sp. nov., 59 & pl. iv. Crag fr. Ramsholt to, 327, 329 fig. ; .... attenuata, sp. nov., 58 & pl. iv. Crag at, 338, 339. -- costatula, 56 & pl. iii ; car. dubia Alderbury Hill (Wilts), flint from, nov., 57 & pl. iii. exhib., cvi ; plateau-gravel at, 297. elegan~ula, sp. nov., 62 & pl. iv. Aldworth (Oxon), Quartzite-gravel nr., elongata, 54 & pl. iii ; vat. cingu- 591. lata nov., 55 & pl. iii; vat. varians Alexandra Land (Franz Josef Land), nov., 55 & pl. iii. geol. feats, of, 631. -- grantoneusis, sp. nov., 60 & pl. iv. hlgous (?) borings, infilled w. silicate ---parwda, sp. nov., 64 & pl. v. of iron, 322-323 fig. -- pulchra, vat. tenuis, 52 & pl. iii ; ALLrORV, S., obituary of, Ix. car. iutermedia nov., 53 & pl. iii. Almannagja (Iceland) basalt, chem. pusilla, sp. nov., 63 & pl. iv. anal. of, 647. quadrata, sp. nov., 61 & pl. iv; Amaltheidm, short period of domin- car. striatissima nov., 62. ance, 448 ; list of genera, 459. similis, sp. nov., 57 & pls. iii-iv. Amlweh (Anglesey), metamorph, grits -- (~.) sulcatula, 64 & pLv. & shales S.W. of, 374-381 figs. ~enuisgriata, sp. nov., 60 & pl. iv. Ammonite-families, & their conn. w. -- terebra, sp. nov., 63 & pl. iv, ' Jurassic' time-divs., 443-444 ; Aclisoides, section of Murchisonia, 66. -

First Three-Dimensionally Preserved in Situ Record of an Aptychophoran Ammonite Jaw Apparatus in the Jurassic and Discussion of the Function of Aptychi
Berliner paläobiologische Abhandlungen 10 321-330 Berlin 2009-11-11 First three-dimensionally preserved in situ record of an aptychophoran ammonite jaw apparatus in the Jurassic and discussion of the function of aptychi Günter Schweigert Abstract: A unique specimen of the microconch ammonite Lingulaticeras planulatum Berckhemer in Ziegler, 1958 comes from a tempestite bed within the Upper Jurassic lithographic limestones of Scham- haupten in Franconia (Painten Formation, uppermost Kimmeridgian). The shell is unique because it retains the complete jaw apparatus in the body chamber. The articulation of the Lamellaptychus and the corresponding upper beak are well preserved. The function of the aptychus is discussed in general, and an operculum function is thought to be unlikely. The formation of strongly calcified aptychi in aspidoceratids and some oppeliid ammonoids is interpreted as an added ballast weight to stabilize the conch for swimming in the water column. Keywords: Ammonites, aptychus, preservation, functional morphology, Upper Jurassic, lithographic lime- stones, Franconia, Germany Zusammenfassung: Ein einzigartig erhaltenes Exemplar des mikroconchen Ammoniten Lingulaticeras planulatum Berckhemer in Ziegler, 1958 aus einer Tempestitlage des oberjurassischen Plattenkalks von Schamhaupten in Franken (Painten-Formation, oberstes Kimmeridgium) enthält noch den vollständigen Kieferapparat in seiner Wohnkammer.Es zeigt die perfekte Artikualation des Lamellaptychus mit dem dazu- gehörenden Oberkiefer. Die Funktion des Aptychus wird allgemein diskutiert und eine Deckelfunktion für unwahrscheinlich gehalten. Die Ausbildung stark verkalkter Aptychen wie in Aspidoceraten und manchen Oppeliiden wird als zusätzliches Tariergewicht gedeutet, um das Gehäuse in starker bewegtem Wasser zu stabilisieren. Schlüsselwörter: Ammoniten, Aptychus, Erhaltung, Plattenkalke, Funktionsmorphologie, Oberjura, Franken, Deutschland Address of the author: Dr. Günter Schweigert, Staatliches Museum für Naturkunde, Rosenstein 1, D-70191 Stuttgart. -

(Oxfordian) Bentonite Deposits in the Paris Basin and the Subalpine Basin, France
Sedimentology (2003) 50, 1035–1060 doi: 10.1046/j.1365-3091.2003.00592.x Characterization and correlation of Upper Jurassic (Oxfordian) bentonite deposits in the Paris Basin and the Subalpine Basin, France PIERRE PELLENARD*, JEAN-FRANCOIS DECONINCK*, WARREN D. HUFF , JACQUES THIERRYà, DIDIER MARCHANDà, DOMINIQUE FORTWENGLER§ and ALAIN TROUILLER– *Morphodynamique continentale et coˆtie`re, UMR 6143 CNRS, University of Rouen, Department of Earth Sciences, 76821 Mont St Aignan Cedex, France (E-mail: [email protected]) University of Cincinnati, Department of Geology, PO Box 210013, Cincinnati, OH 45221-00, USA àBioge´osciences-Dijon, UMR 5561CNRS, Department of Earth Sciences, University of Burgundy, 6 bd Gabriel, 21000 Dijon, France §Le Clos des Vignes, Quartier Perry, 26160 La Be´gude de Mazenc, France –ANDRA, Parc de la Croix Blanche, 1–7 rue Jean Monnet, 92298 Chaˆtenay-Malabry, France ABSTRACT Explosive volcanic activity is recorded in the Upper Jurassic of the Paris Basin and the Subalpine Basin of France by the identification of five bentonite horizons. These layers occur in Lower Oxfordian (cordatum ammonite zone) to Middle Oxfordian (plicatilis zone) clays and silty clays deposited in outer platform environments. In the Paris Basin, a thick bentonite (10–15 cm), identified in boreholes and in outcrop, is dominated by dioctahedral smectite (95%) with trace amounts of kaolinite, illite and chlorite. In contrast, five bentonites identified in the Subalpine Basin, where burial diagenesis and fluid circulation were more important, are composed of a mixture of kaolinite and regular or random illite/smectite mixed-layer clays in variable proportions, indicating a K-bentonite. In the Subalpine Basin, a 2–15 cm thick bentonite underlain by a layer affected by sulphate–carbonate mineralization can be correlated over 2000 km2. -

New Data on the Jaw Apparatus of Fossil Cephalopods
New dataon the jaw apparatus of fossil cephalopods YURI D. ZAKHAROV AND TAMAZ A. LOMINADZE \ Zakharov, Yuri D. & Lominadze, Tamaz A. 19830115: New data on the jaw apparatus of fossil LETHAIA cephalopods. Lethaia, Vol. 16, pp. 67-78. Oslo. ISSN 0024-1164. A newly discovered fossil cephalopod jaw apparatus that may belong to Permian representatives of the Endocochlia is described. Permorhynchus dentatus n. gen. n. sp. is established on the basis of this ~ apparatus. The asymmetry of jaws in the Ectocochlia may be connected with the double function of the ventral jaw apparatus, and the well-developed, relatively large frontal plate of the ventral jaw should be regarded as a feature common to all representatives of ectocochlian cephalopods evolved from early Palaeozoic stock. Distinct features seen in the jaw apparatus of Upper Pcrmian cndocochlians include the pronounced beak form of both jaws and the presence of oblong wings on the ventral mandible. o Cephalopoda. jaw. operculum. aptychus, anaptychus, Permorhynchus n.gen.• evolution. Permian. Yuri D. Zakharov llOpllll Ilscumpueeu« Gaxapoe], Institute of Biology and Pedology, Far-Eastern Scientific Centre. USSR Academy of Science, Vladivostok 690022, USSR (EUOJl020-n9~BnlHbliiuucmu my m Ilaot.neeocmo-cnoro Ha."~H020 uenmpav Axaoeuuu 'HayK CCCP, Bnaoueocmo« 690022, CCCP); Tamaz A. Lominadze ITa.'W3 Apl.j1L10BUl.j Jlouunaoee), Institute of Palaeobiology of Georgian SSR Academy of Science. Tbilisi 380004. USSR (Hncmumvm naJle06UOJl02UU Atcaoeuuu naytc TpY3UHUjKOii CCP. T6'LJUCU 380004. CCP; 19th August. 1980 (revised 1982 06 28). The jaw apparatus of Recent cephalopods is re Turek 1978, Fig. 7, but not the reconstruction in presented by two jaw elements (Fig. -

Maquetación 1
ISSN (print): 1698-6180. ISSN (online): 1886-7995 www.ucm.es /info/estratig/journal.htm Journal of Iberian Geology 33 (2) 2007: 261-282 Callovian and the Callovian - Oxfordian transition sedimentary record in NE Iberian Chain: Taphonomic analysis and palaeogeography El Calloviense y el registro sedimentario del tránsito Calloviense-Oxfordiense en la Cordillera Ibérica nororiental: Análisis tafonómico y paleogeografía G. Meléndez1, J. Ramajo2, J. Bello1, K.N. Page3 1Departamento de Geología (Paleontología). Universidad de Zaragoza. C./ Pedro Cerbuna 12; 50009- Zaragoza (España). [email protected], [email protected] 2Departamento de Geología (Estratigrafía). Universidad de Zaragoza. C./ Pedro Cerbuna 12; 50009- Zaragoza (España). [email protected] 3School of Earth, Ocean & Environmental Sciences, University of Plymouth, Drake Circus, Plymouth PL4 8AA, UK. [email protected] Received: 07/10/06 / Accepted: 30/11/06 Abstract A comparative study is made on Callovian to middle Oxfordian sequences along the Northeastern Iberian Chain (E. Spain). In the NW areas, from Veruela-Ricla to Tosos (SW-S of Zaragoza), Callovian deposits are represented by expanded carbonate sequences (mudstone and marls with a variable content in clastics) ranging from early Bullatus to Athleta Biozone. Farther East in the near area of Moneva-Ariño (Sierra de Arcos) and in the area of Calanda (SE of Zaragoza), this stratigraphic interval is represented by a strongly condensed, 2-4 m thick, highly incomplete sequence. Bullatus to early Anceps biozones are partly represented under bioclastic or oolitic packstone facies. The Callovian-Oxfordian transition interval deposits are represented by a decimeter-thick iron- ooid fossiliferous limestone condensed sequence (low values of sedimentation rate) formed by expanded sediments (high values of instant rate of sediment accumulation). -
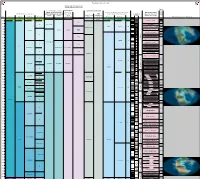
Expanded Jurassic Timescale
TimeScale Creator 2012 chart Russian and Ural regional units Russia Platform regional units Calca Jur-Cret boundary regional Russia Platform East Asian regional units reous stages - British and Boreal Stages (Jur- Australia and New Zealand regional units Marine Macrofossils Nann Standard Chronostratigraphy British regional Boreal regional Cret, Perm- Japan New Zealand Chronostratigraphy Geomagnetic (Mesozoic-Paleozoic) ofossil stages stages Carb & South China (Neogene & Polarity Tethyan Ammonoids s Ma Period Epoch Age/Stage Substage Cambrian) stages Cret) NZ Series NZ Stages Global Reconstructions (R. Blakey) Ryazanian Ryazanian Ryazanian [ no stages M17 CC2 Cretaceous Early Berriasian E Kochian Taitai Um designated ] M18 CC1 145 Berriasella jacobi M19 NJT1 Late M20 7b 146 Lt Portlandian M21 Durangites NJT1 M22 7a 147 Oteke Puaroan Op M22A Micracanthoceras microcanthum NJT1 Penglaizhenian M23 6b 148 Micracanthoceras ponti / Volgian Volgian Middle M24 Burckhardticeras peroni NJT1 Tithonian M24A 6a 149 M24B Semiformiceras fallauxi NJT15 M25 b E 150 M25A Semiformiceras semiforme NJT1 5a Early M26 Semiformiceras darwini 151 lt-Oxf N M-Sequence Hybonoticeras hybonotum 152 Ohauan Ko lt-Oxf R Kimmeridgian Hybonoticeras beckeri 153 m- Lt Late Oxf N Aulacostephanus eudoxus 154 m- NJT14 Late Aspidoceras acanthicum Oxf R Kimmeridgian Kimmeridgian Kimmeridgian Crussoliceras divisum 155 155.431 Card- N Ataxioceras hypselocyclum 156 E Early e-Oxf Sutneria platynota R Idoceras planula Suiningian 157 Cal- Oxf N Epipeltoceras bimammatum 158 lt- Lt Callo -

Text of Draft
Reconnaissance bedrock geologic map for the northern Alaska Peninsula area, southwest Alaska Including the Dillingham, Iliamna, Lake Clark, Taylor Mountains and the western part of the Kenai and Seldovia 1:250,000-scale quadrangles Compiled by Frederic H. Wilson, Robert B. Blodgett, Charles D. Blomé, Solmaz Mohadjer, Cindi C. Preller, Edward P. Klimasauskas, Bruce M. Gamble, and Warren L. Coonrad DISCLAIMER This report is preliminary and has not been reviewed for conformity with U.S. Geological Survey editorial standards or with the North American Stratigraphic Code. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government. This World-Wide-Web publication was prepared by an agency of the United States Government. Neither the United States Government nor any agency thereof, nor any of their employees, makes any warranty, expressed or implied, or assumes any legal liability or responsibility for the accuracy, completeness, or usefulness of any information, apparatus, product, or process disclosed in this report, or represents that its use would not infringe privately owned rights. Reference therein to any specific commercial product, process, or service by trade name, trademark, manufacturer, or otherwise does not necessarily constitute or imply its endorsement, recommendation, or favoring by the United States Government or any agency thereof. Although all data and software published on this Web-site have been used by the USGS, no warranty, expressed or implied, is made by the USGS as to the accuracy of the data and related materials and (or) the functioning of the software. The act of distribution shall not constitute any such warranty, and no responsibility is assumed by the USGS in the use of this data, software, or related materials. -

A Review of the Classification of Jurassic Aspidoceratid Ammonites – the Superfamily Aspidoceratoidea
VOLUMINA JURASSICA, 2020, XVIII (1): 47–52 DOI: 10.7306/VJ.18.4 A review of the classification of Jurassic aspidoceratid ammonites – the Superfamily Aspidoceratoidea Horacio PARENT1, Günter SCHWEIGERT2, Armin SCHERZINGER3 Key words: Superfamily Aspidoceratoidea, Aspidoceratidae, Epipeltoceratinae emended, Peltoceratidae, Gregoryceratinae nov. subfam. Abstract. The aspidoceratid ammonites have been traditionally included in the superfamily Perisphinctoidea. However, the basis of this is unclear for they bear unique combinations of characters unknown in typical perisphinctoids: (1) the distinct laevaptychus, (2) stout shells with high growth rate of the whorl section area, (3) prominent ornamentation with tubercles, spines and strong growth lines running in parallel over strong ribs, (4) lack of constrictions, (5) short to very short bodychamber, and (6) sexual dimorphism characterized by minia- turized microconchs and small-sized macroconchs besides the larger ones, including changes of sex during ontogeny in many cases. Considering the uniqueness of these characters we propose herein to raise the family Aspidoceratidae to the rank of a superfamily Aspi- doceratoidea, ranging from the earliest Late Callovian to the Early Berriasian Jacobi Zone. The new superfamily includes two families, Aspidoceratidae (Aspidoceratinae, Euaspidoceratinae, Epipeltoceratinae and Hybonoticeratinae), and Peltoceratidae (Peltoceratinae and Gregoryceratinae nov. subfam.). The highly differentiated features of the aspidoceratoids indicate that their life-histories -

Abhandlungen Der Geologischen Bundesanstalt in Wien
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Abhandlungen der Geologischen Bundesanstalt in Wien Jahr/Year: 2002 Band/Volume: 57 Autor(en)/Author(s): Sprey Anton Martin Artikel/Article: Early Ontogeny of three Callovian Ammonite Genera (Binatisphinctes, Kosmoceras (Spinikosmoceras) and Hecticoceras) from Ryazan (Russia) 225-255 ©Geol. Bundesanstalt, Wien; download unter www.geologie.ac.at ABHANDLUNGEN DER GEOLOGISCHEN BUNDESANSTALT Abh. Geol. B.-A. ISSN 0016–7800 ISBN 3-85316-14-X Band 57 S. 225–255 Wien, Februar 2002 Cephalopods – Present and Past Editors: H. Summesberger, K. Histon & A. Daurer Early Ontogeny of three Callovian Ammonite Genera (Binatisphinctes, Kosmoceras (Spinikosmoceras) and Hecticoceras) from Ryazan (Russia) ANTON MARTIN SPREY*) 15 Text-Figures, 3 Tables and 8 Plates Callovian Ammonoidea Shell Structure Early Ontogeny Micro-ornament Contents Zusammenfassung ...................................................................................................... 225 Abstract ................................................................................................................. 226 1. Introduction ............................................................................................................. 226 2. Material and Methods .................................................................................................... 226 2.1. Examined Taxa and Their Source ................................................................................... -

GEOS 33000/EVOL 33000: Winter 2006 Course Outline Page 1
GEOS 33000/EVOL 33000: Winter 2006 Course Outline Page 1 Meetings: Tuesdays and Thursdays, 10:30-12:00, HGS 180 Nature of the course: This course is an introduction to mathematical modeling as applied to problems in paleobiology and evolutionary biology. Topics include: basic probability theory; general approaches to modeling; model comparison using likelihood and other criteria; forward modeling of branching processes; sampling models; and inverse methods. A series of programming exercises and a term project are required. Programming in R or C is recommended, but any language may be used. Requirements: 1. Short exercises 2. Readings to be assigned 3. Term paper: Original research, relevant to topics covered in course. Publication length and quality. Brief presentation of research. Prerequisites: Mathematics through first-year calculus; basic computer programming skills (or willingness to learn) URL for course materials: http://geosci.uchicago.edu/∼foote/MODEL Reading General Reference Works Burnham, K.P., and D. R. Anderson. 1998. Model selection and inference: a practical information-theoretic approach. Springer, New York. Edwards, A.W.F. 1992. Likelihood. Johns Hopkins U. Press. Efron, B. and R.J. Tibshirani. 1993. An introduction to the bootstrap. Chapman and Hall, N.Y. Feller, W. 1968 (and other editions). An introduction to probability theory and its applications (two volumes). John Wiley and Sons, N.Y. Gilinsky, N.L., and P.W. Signor, eds. 1991. Analytical paleobiology. Short Courses in Paleontology Number 4. Paleontological Society, Knoxville, Tenn. Gumbel, E.J. 1958. Statistics of extremes. Columbia University Press, N.Y. Press, W.H., S.A. Teukolsky, W.T. Vetterling, B.P. -

Revize Jurské Amonitové Fauny Z Moravského Krasu a Brna Diplomová Práce
PŘÍRODOVĚDECKÁ FAKULTA Revize jurské amonitové fauny z Moravského krasu a Brna Diplomová práce Bc. Petr Hykš Vedoucí práce: Mgr. Tomáš Kumpan, Ph.D. Ústav geologických věd obor Geologie Brno 2020 Bibliografický záznam Autor: Bc. Petr Hykš Přírodovědecká fakulta Masarykova univerzita Ústav geologických věd Název práce: Revize jurské amonitové fauny z Moravského krasu a Brna Studijní program: PřF N-GE Geologie, magisterský studijní program Studijní obor: Geologie Vedoucí práce: Mgr. Tomáš Kumpan, Ph.D. Rok: 2020 Počet stran: 70+15 Klíčová slova: Český masiv, callov, oxford, amoniti, taxonomie, biostratigrafie, provincialismus, paleogeografie Bibliographic record Author: Bc. Petr Hykš Faculty of Science Masaryk University Department of Geological Sciences Title of Thesis: Revision of Jurassic ammonite fauna from the Moravian Karst and Brno Degree Programme: PřF N-GE Geology, Master's degree programme Field of Study: Geology Supervisor: Mgr. Tomáš Kumpan, Ph.D. Year: 2020 Number of Pages: 70+15 Keywords: Bohemian Massif, Callovian, Oxfordian, ammonites, taxonomy, biostratigraphy, provincialism, paleogeography Abstrakt Diplomová práce je zaměřena na taxonomickou revizi středně a pozdně jurských (callov-oxford) amonitů (Ammonitida), skupinu vymřelých hlavonožců z řad amonoidů (Ammonoidea). Studováni byli především amoniti z jurských vápenců na lokalitách Brno-Hády a Olomučany. Přesné stáří jurských vrstev na těchto lokalitách, především v měřítku amonitových zón a subzón, nebylo doposud jednoznačně určeno. Vzhledem k tomu, že amoniti z těchto lokalit byli zpracovávaní naposledy před mnoha dekádami a poznání jejich taxonomie a biostratigrafie vyznamně pokročilo, vyvstala potřeba provést revizi. Na předešlé výzkumy bylo navázáno moderním zpracováním amonitů zohledňujícím vnitrodruhovou variaci a sexuální dimorfismus. Soubor studovaných amonitů je tvořen z části historickými nálezy uloženými v univerzitních a muzejních sbírkách a z části soudobými nálezy autora, jeho kolegů a místních sběratelů.