Root Formation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Ono -- PTH-Pthrp Receptor Signaling in Osterix-Expressing Progenitors.Pdf
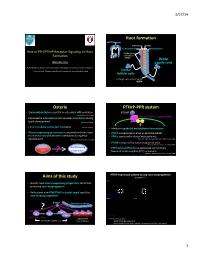
3/27/14 Root forma)on Cementum Dentin Cementoblast Odontoblast Role of PTH/PTHrP Receptor Signaling on Root Epithelial rests Formaon of Malassez (ERM) Dental Wanida Ono papilla cells AGE Orthodon;cs, Department of Developmental Biology, Harvard School of Dental Medicine Endocrine Unit, MassachuseLs General Hospital and Harvard Medical School Dental follicle cells Hertwig’s epithelial root sheath (HERS) Osterix PTHrP-PPR system • Transcripon factor essen;al to osteoblast differen;aon PTHrP (Nakashima K et al. 2002) PTH/PTHrP receptor (PPR) • Expressed in odontoblasts and alveolar osteoblasts during Gαs Gq tooth development (Chen S et al. 2009) • Controls cellular cementum formaon (Cao Z et al. 2012) • Mediates epithelial-mesenchymal interacons • Osterix-expressing precursors in the perichondrium move • PTHrP is expressed in enamel epithelia/HERS? to bone marrow and become osteoblasts during fetal • PPR is expressed in dental mesenchymes development (Maes C, Kronenberg HM et al. 2010) (Beck et al 1995; Lee Deeds and Segre 1995; Liu et al 1998) • PTHrP is required for tooth erup;on in mice (Philbrick WM, Karaplis AC et al. PNAS 1998) Osterix+ Root-forming • PPR haploinsufficiency is associated with primary cells progenitors failure of tooth erup;on (PFE) in humans ? (Decker E, Weber BH et al. Am J Hum Gen 2008) PTHrP expression paern during root morphogenesis Aims of this study PTHrPLacZ/+ x40 P7 P14 P49 • Iden;fy how osterix-expressing progenitors contribute to murine root morphogenesis • Understand how PTH/PTHrP receptor signal regulates root-forming progenitors PTHrP-LacZ x200 P7 x400 P14 ? PTHrP Epithelial root sheath PPR PTHrP-LacZ Osx+ progenitors Odontoblast PTHrP par;cipates in ……. -

Experimental Induction of Odontoblast Differentiation and Stimulation During Preparative Processes
Cells and Materials Volume 3 Number 2 Article 8 1993 Experimental Induction of Odontoblast Differentiation and Stimulation During Preparative Processes H. Lesot Institut de Biologie Médicale C. Begue-Kirn Institut de Biologie Médicale M. D. Kubler Institut de Biologie Médicale J. M. Meyer Institut de Biologie Médicale A. J. Smith Dental School, Birmingham See next page for additional authors Follow this and additional works at: https://digitalcommons.usu.edu/cellsandmaterials Part of the Biomedical Engineering and Bioengineering Commons Recommended Citation Lesot, H.; Begue-Kirn, C.; Kubler, M. D.; Meyer, J. M.; Smith, A. J.; Cassidy, N.; and Ruch, J. V. (1993) "Experimental Induction of Odontoblast Differentiation and Stimulation During Preparative Processes," Cells and Materials: Vol. 3 : No. 2 , Article 8. Available at: https://digitalcommons.usu.edu/cellsandmaterials/vol3/iss2/8 This Article is brought to you for free and open access by the Western Dairy Center at DigitalCommons@USU. It has been accepted for inclusion in Cells and Materials by an authorized administrator of DigitalCommons@USU. For more information, please contact [email protected]. Experimental Induction of Odontoblast Differentiation and Stimulation During Preparative Processes Authors H. Lesot, C. Begue-Kirn, M. D. Kubler, J. M. Meyer, A. J. Smith, N. Cassidy, and J. V. Ruch This article is available in Cells and Materials: https://digitalcommons.usu.edu/cellsandmaterials/vol3/iss2/8 Cells and Materials, Vol. 3, No. 2, 1993 (Pages201-217) 1051-6794/93$5. 00 +. 00 Scanning Microscopy International, Chicago (AMF O'Hare), IL 60666 USA EXPERIMENTAL INDUCTION OF ODONTOBLAST DIFFERENTIATION AND STIMULATION DURING REPARATIVE PROCESSES 1 1 1 2 2 1 H. -

Journal of Dental Research
Journal of Dental Research http://jdr.sagepub.com/ Cell Differentiation and Matrix Organization in Engineered Teeth A. Nait Lechguer, M.L. Couble, N. Labert, S. Kuchler-Bopp, L. Keller, H. Magloire, F. Bleicher and H. Lesot J DENT RES 2011 90: 583 originally published online 4 February 2011 DOI: 10.1177/0022034510391796 The online version of this article can be found at: http://jdr.sagepub.com/content/90/5/583 Published by: http://www.sagepublications.com On behalf of: International and American Associations for Dental Research Additional services and information for Journal of Dental Research can be found at: Email Alerts: http://jdr.sagepub.com/cgi/alerts Subscriptions: http://jdr.sagepub.com/subscriptions Reprints: http://www.sagepub.com/journalsReprints.nav Permissions: http://www.sagepub.com/journalsPermissions.nav >> Version of Record - Apr 13, 2011 OnlineFirst Version of Record - Feb 4, 2011 What is This? Downloaded from jdr.sagepub.com at Service Commun de la Documentation Université de Strasbourg on September 6, 2013 For personal use only. No other uses without permission. © 2011 International & American Associations for Dental Research RESEARCH REPORTS Biomaterials & Bioengineering A. Nait Lechguer1,2, M.L. Couble3,4, N. Labert3,4, S. Kuchler-Bopp1,2, Cell Differentiation and L. Keller1,2, H. Magloire3,4, F. Bleicher3,4, Matrix Organization in and H. Lesot1,2* Engineered Teeth 1INSERM UMR 977, Faculté de Médecine, 11, rue Humann, F-67085 Strasbourg, France; 2Dental School, University of Strasbourg, Strasbourg, France; 3Université de Lyon, Faculté d’Odontologie, Rue Guillaume Paradin, F-69372 Lyon Cedex 08, France; and 4IGFL, CNRS UMR 5242, Ecole Normale Supérieure, 46 Allée d’Italie, 69364, Lyon Cedex 08, France; *corresponding author, [email protected] J Dent Res 90(5):583-589, 2011 ABSTRACT InTRODuCTIOn Embryonic dental cells were used to check a series of criteria to be achieved for tooth engineering. -

6 Development of the Teeth: Root and Supporting Structures Nagat M
AVERY Chap.06 27-11-2002 10:09 Pagina 108 108 II Development of the Teeth and Supporting Structures 6 Development of the Teeth: Root and Supporting Structures Nagat M. ElNesr and James K. Avery Chapter Outline Introduction Introduction... 108 Objectives... 108 Root development is initiated through the contributions Root Sheath Development... 109 of the cells originating from the enamel organ, dental Single-Root Formation... 110 papilla, and dental follicle. The cells of the outer enamel Multiple-Root Formation... 111 epithelium contact the inner enamel epithelium at the Root Formation Anomalies... 112 base of the enamel organ, the cervical loop (Figs. 6.1 and Fate of the Epithelial Root Sheath (Hertwig's Sheath)... 113 6.2A). Later, with crown completion, the cells of the cer- Dental Follicle... 114 vical loop continue to grow away from the crown and Development of (Intermediate) Cementum... 116 become root sheath cells (Figs. 6.2B and 6.3). The inner Cellular and Acellular Cementum... 116 root sheath cells cause root formation by inducing the Development of the Periodontal Ligament... 117 adjacent cells of the dental papilla to become odonto- Development of the Alveolar Process... 119 blasts, which in turn will form root dentin. The root Summary... 121 sheath will further dictate whether the tooth will have Self-Evaluation Review... 122 single or multiple roots. The remainder of the cells of the dental papilla will then become the cells of the root pulp.The third compo- nent in root formation, the dental follicle, is the tissue that surrounds the enamel organ, the dental papilla, and the root. -

Oral Histology Dentinogenesis
Lecture 3 Oral Histology Dr. Lubna Dentinogenesis Dentin formation (Dentinogenesis): Begins at the bell stage development in the papillary tissue adjacent to the concave tip of the folded inner enamel epithelium the site where cuspal development begins. From that point, dentin formation spreads down the cusp slope as far as the cervical loop of the enamel organ, and the dentin thickens until all the coronal dentin is formed. In multicusped teeth, dentin formation begins independently at the sites of each future cusp tip and again spreads down the flanks of the cusp slopes until fusion with adjacent formative centers occurs. Dentin thus formed constitutes the dentin of the crown of the tooth, or coronal dentin. Odontoblasts Differentiation:- The differentiation odontoblasts from the dental papilla in normal development is brought about by the expression of signaling molecules and growth factors in the cells of the inner enamel epithelium illustrate the differentiation sequence, The few organelles. At this time they are separated from the inner enamel epithelium by cell free zone that contains some fine collagen fibrils. Almost immediately after cells of the inner enamel epithelium reverse polarity, changes also occur in the adjacent dental papilla. The ectomesenchymal cells adjoining this zone rapidly enlarge and elongate to become preodontoblasts first and then odontoblasts as their cytoplasm increases in volume to contain increasing amounts of protein-synthesizing organelles. Then this zone gradually is eliminated as the odontoblasts differentiate and increase in size and occupy this zone. These newly differentiated cells are characterized by being highly polarized (40 µm in length and 7 µm in width), with their nuclei positioned away from the inner enamel epithelium. -

Dental Cementum Reviewed: Development, Structure, Composition, Regeneration and Potential Functions
Braz J Oral Sci. January/March 2005 - Vol.4 - Number 12 Dental cementum reviewed: development, structure, composition, regeneration and potential functions Patricia Furtado Gonçalves 1 Enilson Antonio Sallum 1 Abstract Antonio Wilson Sallum 1 This article reviews developmental and structural characteristics of Márcio Zaffalon Casati 1 cementum, a unique avascular mineralized tissue covering the root Sérgio de Toledo 1 surface that forms the interface between root dentin and periodontal Francisco Humberto Nociti Junior 1 ligament. Besides describing the types of cementum and 1 Dept. of Prosthodontics and Periodontics, cementogenesis, attention is given to recent advances in scientific Division of Periodontics, School of Dentistry understanding of the molecular and cellular aspects of the formation at Piracicaba - UNICAMP, Piracicaba, São and regeneration of cementum. The understanding of the mechanisms Paulo, Brazil. involved in the dynamic of this tissue should allow for the development of new treatment strategies concerning the approach of the root surface affected by periodontal disease and periodontal regeneration techniques. Received for publication: October 01, 2004 Key Words: Accepted: December 17, 2004 dental cementum, review Correspondence to: Francisco H. Nociti Jr. Av. Limeira 901 - Caixa Postal: 052 - CEP: 13414-903 - Piracicaba - S.P. - Brazil Tel: ++ 55 19 34125298 Fax: ++ 55 19 3412 5218 E-mail: [email protected] 651 Braz J Oral Sci. 4(12): 651-658 Dental cementum reviewed: development, structure, composition, regeneration and potential functions Introduction junction (Figure 1). The areas and location of acellular Cementum is an avascular mineralized tissue covering the afibrillar cementum vary from tooth to tooth and along the entire root surface. Due to its intermediary position, forming cementoenamel junction of the same tooth6-9. -

Basic Histology (23 Questions): Oral Histology (16 Questions
Board Question Breakdown (Anatomic Sciences section) The Anatomic Sciences portion of part I of the Dental Board exams consists of 100 test items. They are broken up into the following distribution: Gross Anatomy (50 questions): Head - 28 questions broken down in this fashion: - Oral cavity - 6 questions - Extraoral structures - 12 questions - Osteology - 6 questions - TMJ and muscles of mastication - 4 questions Neck - 5 questions Upper Limb - 3 questions Thoracic cavity - 5 questions Abdominopelvic cavity - 2 questions Neuroanatomy (CNS, ANS +) - 7 questions Basic Histology (23 questions): Ultrastructure (cell organelles) - 4 questions Basic tissues - 4 questions Bone, cartilage & joints - 3 questions Lymphatic & circulatory systems - 3 questions Endocrine system - 2 questions Respiratory system - 1 question Gastrointestinal system - 3 questions Genitouirinary systems - (reproductive & urinary) 2 questions Integument - 1 question Oral Histology (16 questions): Tooth & supporting structures - 9 questions Soft oral tissues (including dentin) - 5 questions Temporomandibular joint - 2 questions Developmental Biology (11 questions): Osteogenesis (bone formation) - 2 questions Tooth development, eruption & movement - 4 questions General embryology - 2 questions 2 National Board Part 1: Review questions for histology/oral histology (Answers follow at the end) 1. Normally most of the circulating white blood cells are a. basophilic leukocytes b. monocytes c. lymphocytes d. eosinophilic leukocytes e. neutrophilic leukocytes 2. Blood platelets are products of a. osteoclasts b. basophils c. red blood cells d. plasma cells e. megakaryocytes 3. Bacteria are frequently ingested by a. neutrophilic leukocytes b. basophilic leukocytes c. mast cells d. small lymphocytes e. fibrocytes 4. It is believed that worn out red cells are normally destroyed in the spleen by a. neutrophils b. -

The Role of Transforming Growth Factor Beta in Tertiary Dentinogenesis
The role of transforming growth factor beta in tertiary dentinogenesis Tetiana Haniastuti1, Phides Nunez2, and Ariadna A. Djais3 1Department of Oral Biology, Gadjah Mada University, Indonesia 2Division of Oral Ecology in Health and Infection, Niigata University, Japan 3Department of Oral Biology, University of Indonesia, Indonesia ABSTRACT The most visible repair response to pulp injury is the deposition of a tertiary dentin matrix over the dentinal tubules of the primary or secondary dentin. Tertiary dentin is distinguished as reactionary and reparative dentin, depending on the severity of the initiating response and the conditions under which the newly deposited dentin matrix was elaborated. Transforming growth factor beta (TGF-b) superfamily is a large group of growth factors that serve important roles in regulating cell growth, differentiation, and function. Members of this superfamily have been implicated in the repair process of the dental tissue after injury. Although numerous studies have proved that those bioactive molecules carry out an important role in the formation of tertiary dentin, comprehensive report regarding that phenomenon is not yet available. This review article aimed to summarize the role of TGF-b on tertiary dentinogenesis during the progression of a carious lesion. Key words: transforming growth factor beta, tertiary dentinogenesis, reactionary dentin, reparative dentin Correspondence: Tetiana Haniastuti, c/o: Bagian Biologi Oral, Fakultas Kedokteran Gigi Universitas Gadjah Mada. Jln. Denta no. 1, Sekip Utara Yogyakarta 55281, Indonesia. E-mail: [email protected] INTRODUCTION such as cell proliferation, differentiation, and matrix synthesis. Recent studies showed that this substance has a The complex structural composition of teeth provides significant role in the immune response7 and tissue repair8 hardness and durability as a barrier against bacterial of the dental pulp. -

Effects of Lipid Metabolism on Mouse Incisor Dentinogenesis
www.nature.com/scientificreports OPEN Efects of lipid metabolism on mouse incisor dentinogenesis Yutaro Kurotaki1,2,3, Nobuhiro Sakai2,3*, Takuro Miyazaki4, Masahiro Hosonuma2,3,5, Yurie Sato2,3,6, Akiko Karakawa2,3, Masahiro Chatani2,3, Mie Myers1, Tetsuo Suzawa7, Takako Negishi-Koga2,3,8, Ryutaro Kamijo7, Akira Miyazaki4, Yasubumi Maruoka1 & Masamichi Takami2,3* Tooth formation can be afected by various factors, such as oral disease, drug administration, and systemic illness, as well as internal conditions including dentin formation. Dyslipidemia is an important lifestyle disease, though the relationship of aberrant lipid metabolism with tooth formation has not been clarifed. This study was performed to examine the efects of dyslipidemia on tooth formation and tooth development. Dyslipidemia was induced in mice by giving a high-fat diet (HFD) for 12 weeks. Additionally, LDL receptor-defcient (Ldlr−/−) strain mice were used to analyze the efects of dyslipidemia and lipid metabolism in greater detail. In the HFD-fed mice, incisor elongation was decreased and pulp was signifcantly narrowed, while histological fndings revealed disappearance of predentin. In Ldlr−/− mice fed regular chow, incisor elongation showed a decreasing trend and pulp a narrowing trend, while predentin changes were unclear. Serum lipid levels were increased in the HFD- fed wild-type (WT) mice, while Ldlr−/− mice given the HFD showed the greatest increase. These results show important efects of lipid metabolism, especially via the LDL receptor, on tooth homeostasis maintenance. In addition, they suggest a diferent mechanism for WT and Ldlr−/− mice, though the LDL receptor pathway may not be the only factor involved. A regular high-fat diet (HFD) has been shown to result in such lifestyle diseases as dyslipidemia, obesity, and diabetes1,2. -

A Global Compendium of Oral Health
A Global Compendium of Oral Health A Global Compendium of Oral Health: Tooth Eruption and Hard Dental Tissue Anomalies Edited by Morenike Oluwatoyin Folayan A Global Compendium of Oral Health: Tooth Eruption and Hard Dental Tissue Anomalies Edited by Morenike Oluwatoyin Folayan This book first published 2019 Cambridge Scholars Publishing Lady Stephenson Library, Newcastle upon Tyne, NE6 2PA, UK British Library Cataloguing in Publication Data A catalogue record for this book is available from the British Library Copyright © 2019 by Morenike Oluwatoyin Folayan and contributors All rights for this book reserved. No part of this book may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, electronic, mechanical, photocopying, recording or otherwise, without the prior permission of the copyright owner. ISBN (10): 1-5275-3691-2 ISBN (13): 978-1-5275-3691-3 TABLE OF CONTENTS Foreword .................................................................................................. viii Introduction ................................................................................................. 1 Dental Development: Anthropological Perspectives ................................. 31 Temitope A. Esan and Lynne A. Schepartz Belarus ....................................................................................................... 48 Natallia Shakavets, Alexander Yatzuk, Klavdia Gorbacheva and Nadezhda Chernyavskaya Bangladesh ............................................................................................... -

Role of Connexin 43 in Odontoblastic Differentiation and Structural Maintenance in Pulp Damage Repair
International Journal of Oral Science www.nature.com/ijos ARTICLE OPEN Role of connexin 43 in odontoblastic differentiation and structural maintenance in pulp damage repair Jiaxin Yin1,2, Jue Xu3, Ran Cheng1, Meiying Shao3, Yuandong Qin1, Hui Yang1 and Tao Hu1 Dental pulp can initiate its damage repair after an injury of the pulp–dentin complex by rearrangement of odontoblasts and formation of newly differentiated odontoblast-like cells. Connexin 43 (Cx43) is one of the gap junction proteins that participates in multiple tissue repair processes. However, the role of Cx43 in the repair of the dental pulp remains unclear. This study aimed to determine the function of Cx43 in the odontoblast arrangement patterns and odontoblastic differentiation. Human teeth for in vitro experiments were acquired, and a pulp injury model in Sprague-Dawley rats was used for in vivo analysis. The odontoblast arrangement pattern and the expression of Cx43 and dentin sialophosphoprotein (DSPP) were assessed. To investigate the function of Cx43 in odontoblastic differentiation, we overexpressed or inhibited Cx43. The results indicated that polarized odontoblasts were arranged along the pulp–dentin interface and had high levels of Cx43 expression in the healthy teeth; however, the odontoblast arrangement pattern was slightly changed concomitant to an increase in the Cx43 expression in the carious teeth. Regularly arranged odontoblast-like cells had high levels of the Cx43 expression during the formation of mature dentin, but the odontoblast- like cells were not regularly arranged beneath immature osteodentin in the pulp injury models. Subsequent in vitro experiments demonstrated that Cx43 is upregulated during odontoblastic differentiation of the dental pulp cells, and inhibition or overexpression of Cx43 influence the odontoblastic differentiation. -

Developmental Biology of Cementum
Int. J. Dev. Biol. 45: 695-706 (2001) Review Developmental Biology of Cementum THOMAS G.H. DIEKWISCH* Allan G. Brodie Laboratory for Craniofacial Genetics, University of Illinois at Chicago, USA CONTENTS Origins of cementum - a scientific "whodunit" ........................................................................695 Loss of ameloblast continuity and insertion of mesenchymal cells from the dental follicle proper ................................................................................................697 Initial cementum matrix deposition by mesenchymal cells in proximity to non-secretory epithelial cells ...................................................................................699 Cementogenesis at the tooth cervix and at the cemento-enamel junction .............................700 Early removal of HERS from the root surface in humans as seen in the Gottlieb collection ..............................................................................................701 Role of amelogenins in cementogenesis ................................................................................702 Possible mechanism of cementoblast induction .....................................................................704 Summary ................................................................................................................................704 KEY WORDS: Cementum, Hertwig’s epithelial root sheath, Gottlieb, amelogenin, periodontium Tooth cementum is a bone-like mineralized tissue secreted by Origins of cementum - a scientific