Backed Woodpecker Status Review
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Wildfire Mitigation Plan
San Diego Gas & Electric Company Wildfire Mitigation Plan February 7, 2020 Table of Contents Executive Summary ....................................................................................................................... viii 1. Persons Responsible for Executing the WMP ......................................................................... 1 1.1. Verification ....................................................................................................................... 3 2. Metrics and Underlying Data .................................................................................................. 4 2.1. Lessons Learned: How Tracking Metrics on the 2019 Plan Has Informed the 2020 Plan ......................................................................................................................... 4 2.2. Recent Performance on Progress Metrics, Last 5 Years .................................................. 7 2.3. Recent Performance on Outcome Metrics, Annual and Normalized for Weather, Last 5 Years ................................................................................................................................. 7 2.4. Description of Additional Metrics .................................................................................... 8 2.5. Description of Program Targets ....................................................................................... 8 2.6. Detailed Information Supporting Outcome Metrics ........................................................ 9 2.7. Mapping Recent, -

Attachment a ‐ Forest Service Wildfire, NEPA, and Salvage Summary
Attachment A ‐ Forest Service Wildfire, NEPA, and Salvage Summary Fiscal Year 2007 2008 2009 2010 2011 2012 2013 2014 2015 2016 2007‐2016 Number of Fires 1 63 64 53 33 66 79 56 56 127 110 707 Total fire acres on NFS 2 1,751,118 1,326,893 549,108 211,327 1,489,029 2,411,413 1,141,353 741,465 1,587,843 1,038,686 12,248,235 High severity acres on NFS 3 842,658 368,595 268,944 76,192 619,020 809,720 513,957 265,045 489,668 397,654 4,651,453 Number of NEPA decisions identified 4 129 Acres of salvage planned in NEPA 5 218 17,255 2,134 14,010 22,761 28,937 13,809 13,264 112,388 Number of NEPA decisions litigated 6 125110332422 Litigation cases won by USFS 7 013110131112 Litigation cases lost by USFS 8 1120001011 7 Litigation cases pending 9 0000001002 3 Acres of salvage reported accomplished 10 328 2,665 8,125 3,464 8,774 6,916 11,672 19,792 16,926 21,234 99,896 1 Fires burning more than 1,000 acres on NFS land 10 Salvage harvest activity records identified as awarded in Forest Service Activity 2 Total acres inside fire perimeter on NFS land Tracking System (FACTS) by GIS analysis of fire perimeters. 3 Classified as greater than 75% mortality using Rapid Assessment of Vegetation Condition after Wildfire (RAVG) 4 Identified by fire salvage keyword search in PALS (Planning Appeals and Disclaimer: Only the litigation data is believed to be 100% complete and Litigation System) or reported with sale activity records in Forest Service systems accurate. -

Office of Governor Kate Brown
23 Oregon’s economy continues to do well. Businesses are growing, unemployment is low, and wages are increasing. However, not all Oregonians are enjoying this prosperity equally. We need to be diligent champions of diversity, equity, and inclusion in our work, in our communities, and in our regions. The foundation of the Regional Solutions program recognizes that Oregon is comprised of many different economies and tailors the state’s support to create thriving communities across the state. Regional Solutions staff live and work in the communities they serve, making sure state agencies work together efficiently and collaborate with local partners. The staff work with a grassroots approach powered by the ability to cross-cut agencies to assist businesses, local governments, and partners to get things done. They work on the nuts and bolts of economic development: streamlining permits, advising on land use, and building partnerships between the private, public, and philanthropic sectors. We see the results when businesses grow and things get built: transportation networks, water systems, broadband, homes, innovation centers, and more. That leads to not just more jobs, but better jobs across the state. This is how we support sustained growth rooted in our local communities and their plans to support economic vitality. With the impressive bench strength of the Regional Solutions staff, I give special assignments that move the needle for initiatives of state wide significance. In 2018, Regional Solutions took on workforce housing. Today, we are partnering in communities across the state on five housing pilots that will inform solutions that innovatively address this important issue. -

Recollections of Initial Attack of the Barry Point Fire, the First Three Days
Recollections of initial attack of the Barry Point Fire, the first three days Meeting Date: Sept 11, 2012 Location: Lakeview Interagency office Gearhart conference room Reason for Meeting: Fred Way, Fremont-Winema Forest Supervisor, requested information on the beginning days of the Barry Point Fire to get an understanding of how the Forest responded. Note Taker: Lucinda Nolan and Glen Westlund, Forest Environmental Coordinator, FOIA coordinator, and litigation coordinator Attending: Fred Way, Forest Supervisor; Coley Neider, 8/6 Operations, Trent Wilkie, 8/6 Division A Supervisor, Chad Bergren, 8/6 Division B Supervisor, Clark Hammond, air operations (on phone), Helitak Foreman; Dustin Gustaveson, Oregon Dept of Forestry Supervisor (not assigned to fire); Barry Shullanberger, 8/7-8/8 Day Operations; Noel Livingston, Not present during incident, currently acting Fremont-Winema NF Fire Staff officer; Bob Crumrine, Federal Duty Officer; Eric Knerr, 8/6-8/7 (morning) IC and logistical support; Mitch Wilson, 8/7-8/8 Division Z Supervisor; Abel Harrington, 8/7-8/8 Division A Supervisor; Kevin Burdon, Not assigned to fire ODF Supervisor. Evening August 5: Lighting storm begins with the first strike occurring at 8:59 am (0859) and the last strike at 10:56 pm (2256). See Attachment 1, Lightning Display Map. Day of August 6th 2012 Due to lightning incidents on 8/5 and expecting more on 8/6 the Lakeview Interagency Fire Center is placed in lightning mode (see attachment 2 and 3 for roles and responsibilities) which dispatches two resources per event as they get reported. The Forest has seven engines and many lookouts (5) manned (in service) by 0700, in preparation for any potential fires from previous night’s storm. -

Safety Element
TABLE OF CONTENTS Chapter 6: Safety Element 1. INTRODUCTION ........................................................................................................................................ 1 PURPOSE, SCOPE, AND CONTENT ...................................................................................................................... 1 REGULATORY FRAMEWORK ............................................................................................................................... 2 RELATIONSHIP TO OTHER DOCUMENTS .............................................................................................................. 3 Other General Plan Elements .................................................................................................................... 3 Area Plans .................................................................................................................................................. 3 Multi-Jurisdictional Local Hazard Mitigation Plan ....................................................................................... 3 RELATIONSHIP TO THE EMERGENCY MANAGEMENT DEPARTMENT ....................................................................... 4 HAZARD REDUCTION ......................................................................................................................................... 4 2. ISSUES AND POLICIES ............................................................................................................................ 6 CODE CONFORMANCE AND DEVELOPMENT REGULATIONS -
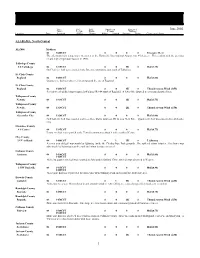
Storm Data and Unusual Weather Phenomena
Storm Data and Unusual Weather Phenomena Time Path Path Number of Estimated June 2002 Local/ Length Width Persons Damage Location Date Standard (Miles) (Yards) Killed Injured Property Crops Character of Storm ALABAMA, North Central ALZ006 Madison 03 1600CST 0 0 0 0 Excessive Heat The afternoon high temperature measured at the Huntsville International Airport was 95 degrees. This reading tied the previous record high temperature last set in 1998. Talladega County 5 S Talladega 04 1219CST 0 0 5K 0 Hail(1.75) Golf ball size hail was reported in the Brecon community, just south of Talladega. St. Clair County Ragland 04 1301CST 0 0 0 0 Hail(1.00) Quarter size hail was observed in and around the city of Ragland. St. Clair County Ragland 04 1301CST 0 0 3K 0 Thunderstorm Wind (G55) Several trees had their tops snapped off along SR 144 south of Ragland. A few of the downed trees snapped power lines. Tallapoosa County Newsite 04 1314CST 0 0 2K 0 Hail(1.75) Tallapoosa County Newsite 04 1314CST 0 0 2K 0 Thunderstorm Wind (G50) Tallapoosa County Alexander City 04 1320CST 0 0 0 0 Hail(1.00) Golf ball size hail was reported and trees were blown down on SR 22 near New Site. Quarter size hail was observed in Alexander City. Cherokee County 4 S Centre 04 1335CST 0 0 0 0 Hail(0.75) Penny size hail was reported at the Tennala community about 4 miles south of Centre. Clay County 13 N Ashland 04 1400CST 0 1 3K 0 Lightning A seven year old girl was struck by lightning inside the Cheaha State Park grounds. -

Annual Report
RIVERSIDE COUNTY FIRE DEPARTMENT IN COOPERATION WITH CAL FIRE ANNUAL REPORT 2017 MISSION STATEMENT Riverside County Fire Department is a public safety agency dedicated to protecting life, property and the environment through professionalism, integrity and efficiency. I VISION STATEMENT Riverside County Fire Department is committed to exemplary service and will be a leader in Fire protection and emergency services through continuous improvement, innovation and the most efficient and responsible use of resources. II III TABLE OF CONTENTS Mission Statement County Fire Chief’s Message 2 Organizational Structure 4 Response Statistics 9 Administration 18 Air Program 24 Camp Program 28 Communications/Information Technology 32 Emergency Command Center 36 Emergency Medical Services 40 Law Enforcement/Hazard Reduction 44 Fleet Services 48 Health and Safety 50 Office of the Fire Marshal 54 Pre Fire Management 58 Public Affairs Bureau/Education 62 Service Center 66 Strategic Planning 70 Training 74 Volunteer Reserve Program 78 Retirements/In Memoriam 80 The Year in Pictures 82 Acknowledgements 94 IV MESSAGE FROM THE FIRE CHIEF CAL FIRE AND RIVERSIDE COUNTY FIRE CHIEF DANIEL R. TALBOT 2 It is with pride that my staff and I publish this report. I am indeed proud of our service-oriented Fire Department. The combination of the State, County and locally funded fire resources has created a truly integrated, cooperative and regional fire protection system. This system has the capacity to respond to 452 requests for service daily and the resiliency, due to our depth of resources, to simultaneously respond to major structure and wildland fires. In 2017, our Fire Department responded to 164,594 requests for service. -

Wildfire Mitigation Plan
San Diego Gas & Electric Company Wildfire Mitigation Plan February 7, 2020 (Rev 1.) March 2, 2020 Table of Contents Executive Summary .................................................................................................................... viii 1. Persons Responsible for Executing the WMP ........................................................................ 1 1.1. Verification ..................................................................................................................... 3 2. Metrics and Underlying Data ................................................................................................. 4 2.1. Lessons Learned: How Tracking Metrics on the 2019 Plan Has Informed the 2020 Plan ........................................................................................................................ 4 2.2. Recent Performance on Progress Metrics, Last 5 Years .................................................. 7 2.3. Recent Performance on Outcome Metrics, Annual and Normalized for Weather, Last 5 Years ............................................................................................................................... 7 2.4. Description of Additional Metrics ................................................................................... 8 2.5. Description of Program Targets ...................................................................................... 8 2.6. Detailed Information Supporting Outcome Metrics ....................................................... 9 2.7. Mapping -

1 White-Headed Woodpecker Monitoring on the Barry Point Fire
White-headed Woodpecker monitoring on the Barry Point Fire for the Lakeview Stewardship CFLR, Fremont-Winema National Forest, 2013 progress report Finalized February 2014 by: USFS Rocky Mountain Research Station Victoria Saab, Jonathan Dudley, and Quresh Latif To: Fremont-Winema National Forest Amy Markus, Forest Wildlife Biologist Introduction The Collaborative Forest Landscape Restoration (CFLR) program is a cooperative effort to increase the rate of restoration on our National Forests. Monitoring is a key component of the CFLR program and our work is designed to address how well CFLR projects are meeting their forest restoration and wildlife habitat conservation goals. The white-headed woodpecker (Picoides albolarvatus; WHWO) is a regional endemic species of the Inland Northwest and may be particularly vulnerable to environmental change because it occupies a limited distribution and has narrow habitat requirements in dry coniferous forests. Monitoring in CFLR projects, such as the Lakeview Stewardship CFLR project on the Fremont-Winema National Forest (FreWin), also contributes to other ongoing, regional efforts to monitor effectiveness of silvicultural and prescribed-fire treatments for white-headed woodpeckers throughout their range in Oregon, Idaho, and Washington. Vegetation and fuels data collection also support modeling of fire-climate impacts on future forest conditions and wildlife habitat suitability. To meet their various ecological needs, white-headed woodpeckers require heterogeneous landscapes characterized by a mosaic of open- and closed-canopied ponderosa pine forests (Wightman et al. 2010, Hollenbeck et al. 2011), which are expected to benefit vascular plant and other vertebrate wildlife populations (e.g., Noss et al. 2006). Consequently, monitoring white- headed woodpecker populations and their habitat associations is central to biological monitoring for the Lakeview Stewardship CFLR project on the FreWin National Forest. -

11.12.20 Comment Letter Re Otay Ranch Village 13
XAVIER BECERRA State of California Attorney General DEPARTMENT OF JUSTICE 600 WEST BROADWAY, SUITE 1800 SAN DIEGO, CA 92101 P.O. BOX 85266 SAN DIEGO, CA 92186-5266 Public: (619) 738-9000 Telephone: (619) 738-9519 Facsimile: (619) 645-2271 E-Mail: [email protected] November 12, 2020 San Diego County Planning & Development Services Attn: Mark Wardlaw, Director of Planning & Development Services 5510 Overland Avenue, Suite 310 San Diego, CA 92123 By email: [email protected] RE: Otay Ranch Resort Village—Village 13 Final Environmental Impact Report; Otay Ranch Resort Village, Project Nos. GPA04-003, REZ04-009, TM-5361, SP04-002, and ER LOG04-19-005 Dear Mr. Wardlaw: We appreciate your preparation of a Final Environmental Impact Report (FEIR) responding to public comments on the Draft Environmental Impact Report (DEIR), including the comments we submitted on December 27, 2019, regarding wildfire risks associated with the proposed Otay Ranch Resort Village—Village 13 Development (Project). After reviewing the FEIR, we acknowledge and appreciate that you have provided more information regarding wildfire risks associated with the Project. We believe, however, that the FEIR’s discussion of these risks remains inadequate.1 I. THE FEIR FAILS TO ADEQUATELY ADDRESS THE INCREASED WILDFIRE RISK THAT WILL RESULT FROM THE PROJECT In our comment letter, we explained that locating new development in a very high fire hazard severity zone will itself increase the risk of fire and, as a result, increase the risk of exposing residents, employees, and visitors to that enhanced risk. We further explained that the DEIR fails to analyze the increased risk of wildfire that will result from siting the Project within a such a zone. -

2018/2019 Grant Awards
California Climate Investments CAL FIRE Forest Health and Forest Legacy Fiscal Year 2018-2019 Grant Awards Applicant Project Title County(ies) Grant Award Project Description Restore forest health by implementing fuels reduction and hardwood thinning in degraded forests. Pre-commercial and commercial thinning will lead to increased carbon sequestration Bureau of Land Management, Arcata Lacks Creek Management Area Humboldt $ 4,314,243 and support long term forest development in a late successional forest reserve. This is an area Field Office Landscape Restoration threatened by Sudden Oak Death and high fire risk. Project will lead to the development of old growth forest characteristics and improve wildlife habitat and water quality. The Collins Modoc Reforestation Project seeks to reforest approximately 10,143 acres at up to 250 trees per acre of Ponderosa Pine forest lost during the 2012 Barry Point Fire for long-term Collins Timber Company, LLC Collins Modoc Reforestation Project Modoc $ 3,305,164 forest health, sustainable timber harvest, and a potential carbon offset project for the ARB marketplace. The Whiskey CE is Phase 3 of a trio of working forest CEs on the 39,685-acre Scott River Headwaters property (SRH), the largest block of private forestland in Scott Valley. The Whiskey Whiskey Working Forest CE extinguishes development rights on 18,683 acres and helps ensure significant flows of timber Ecotrust Forests II, LLC Conservation Easement - Scott River Siskiyou $ 5,805,798 to nearby mills in perpetuity. SRH is positioned between fire-prone wilderness areas and 5 Headwaters Phase 3 communities. The Whiskey CE and companion CEs will enhance water quality, improve wildlife habitat and forest resiliency, reduce fire risk, and increase carbon storage providing a 1,496,017 CO2-e. -

Unit Identifier Guide: Data Report
A Publication of the National Wildfire Coordinating Group Unit Identifier Guide: Data Report PMS 931 - Published Date: Wednesday, June 17, 2020 Sponsored for NWCG publication by the NWCG Data Management Committee. Prepared and maintained by the Unit Identifier Unit. Questions regarding the content of this product should be directed to the NWCG Unit Identifier Unit (UIU), National Interagency Fire Center, 3833 S Development Avenue, Boise ID 83705, or to the UIU members listed on the NWCG Web site at http://www.nwcg.gov. Questions and comments may also be emailed to [email protected]. This product is available electronically on the NWCG Web site at http://www.nwcg.gov. Previous editions: none. The National Wildfire Coordinating Group (NWCG) has approved the contents of this product for the guidance of its member agencies and is not responsible for the interpretation or use of this information by anyone else. NWCG’s intent is to specifically identify all copyrighted content used in NWCG products. All other NWCG information is in the public domain. Use of public domain information, including copying, is permitted. Use of NWCG information within another document is permitted, if NWCG information is accurately credited to the NWCG. The NWCG logo may not be used except on NWCG authorized information. “National Wildfire Coordinating Group”, “NWCG”, and the NWCG logo are trademarks of the National Wildfire Coordinating Group. The use of trade, firm, or corporation names or trademarks in this product is for the information and convenience of the reader and does not constitute an endorsement by the National Wildfire Coordinating Group or its member agencies of any product or service to the exclusion of others that may be suitable.