Terrestrial Higher Plant Respiration and Net Primary Production I
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Foliar and Ecosystem Respiration in an Old-Growth Tropical Rain Forest
Plant, Cell and Environment (2008) 31, 473–483 doi: 10.1111/j.1365-3040.2008.01775.x Foliar and ecosystem respiration in an old-growth tropical rain forest MOLLY A. CAVALERI1,2, STEVEN F. OBERBAUER3,4 & MICHAEL G. RYAN5,6 1Graduate Degree Program in Ecology, Colorado State University, Fort Collins, CO 80523, USA, 2Botany Department, University of Hawaii, Manoa, 3190 Maile Way, Honolulu, HI 96826, USA, 3Department of Biological Sciences, Florida International University, 11200 SW 8th Street, Miami, FL 33199, USA, 4Fairchild Tropical Botanic Garden, 11935 Old Cutler Road, Miami, FL 33156, USA, 5United States Department of Agriculture Forest Service, Rocky Mountain Research Station, 240 West Prospect RD, Fort Collins, CO 80526, USA and 6Graduate Degree Program in Ecology, Department of Forest, Rangeland, and Watershed Stewardship, Colorado State University, Fort Collins, CO 80523, USA ABSTRACT Chambers et al. 2004). Results vary widely about whether tropical forests are presently acting as carbon sources or Foliar respiration is a major component of ecosystem res- sinks, or how this may be affected by global warming. piration, yet extrapolations are often uncertain in tropical Several eddy flux studies have concluded that tropical rain forests because of indirect estimates of leaf area index forests are primarily acting as carbon sinks (Fan et al. 1990; (LAI). A portable tower was used to directly measure LAI Grace et al. 1995; Malhi et al. 1998; Loescher et al. 2003, and night-time foliar respiration from 52 vertical transects but see Saleska et al. 2003). Many atmosphere–biosphere throughout an old-growth tropical rain forest in Costa Rica. modelling studies, on the other hand, predict that tropical In this study, we (1) explored the effects of structural, func- forests will be an increased carbon source with global tional and environmental variables on foliar respiration; (2) warming (Kindermann, Würth & Kohlmaier 1996; Braswell extrapolated foliar respiration to the ecosystem; and (3) et al. -

7.014 Handout PRODUCTIVITY: the “METABOLISM” of ECOSYSTEMS
7.014 Handout PRODUCTIVITY: THE “METABOLISM” OF ECOSYSTEMS Ecologists use the term “productivity” to refer to the process through which an assemblage of organisms (e.g. a trophic level or ecosystem assimilates carbon. Primary producers (autotrophs) do this through photosynthesis; Secondary producers (heterotrophs) do it through the assimilation of the organic carbon in their food. Remember that all organic carbon in the food web is ultimately derived from primary production. DEFINITIONS Primary Productivity: Rate of conversion of CO2 to organic carbon (photosynthesis) per unit surface area of the earth, expressed either in terns of weight of carbon, or the equivalent calories e.g., g C m-2 year-1 Kcal m-2 year-1 Primary Production: Same as primary productivity, but usually expressed for a whole ecosystem e.g., tons year-1 for a lake, cornfield, forest, etc. NET vs. GROSS: For plants: Some of the organic carbon generated in plants through photosynthesis (using solar energy) is oxidized back to CO2 (releasing energy) through the respiration of the plants – RA. Gross Primary Production: (GPP) = Total amount of CO2 reduced to organic carbon by the plants per unit time Autotrophic Respiration: (RA) = Total amount of organic carbon that is respired (oxidized to CO2) by plants per unit time Net Primary Production (NPP) = GPP – RA The amount of organic carbon produced by plants that is not consumed by their own respiration. It is the increase in the plant biomass in the absence of herbivores. For an entire ecosystem: Some of the NPP of the plants is consumed (and respired) by herbivores and decomposers and oxidized back to CO2 (RH). -

Acclimation of Photosynthesis and Respiration to Elevated Atmospheric CO2 in Two Scrub Oaks
Global Change Biology (2002) 8, 317±328 Acclimation of photosynthesis and respiration to elevated atmospheric CO2 in two Scrub Oaks GRAHAM J. HYMUS, TOM G. SNEAD, DAVID P. JOHNSON, BRUCE A. HUNGATE* and BERT G.DRAKE Smithsonian Environmental Research Center, Mailcode DYN-2, Kennedy Space Center, FL 32899, USA, *Department of Biological Sciences and Merriam-Powell Center for Environmental Research, Box 5640, Northern Arizona University, Flagstaff, AZ 86011- 5640, USA Abstract For two species of oak, we determined whether increasing atmospheric CO2 concen- tration (Ca)would decrease leaf mitochondrial respiration (R)directly, or indirectly owing to their growth in elevated Ca , or both. In particular, we tested whether acclima- tory decreases in leaf-Rubisco content in elevated Ca would decrease R associated with its maintenance. This hypothesis was tested in summer 2000 on sun and shade leaves of Quercus myrtifolia Willd. and Quercus geminata Small. We also measured R on five occasions between summer 1999 and 2000 on leaves of Q. myrtifolia. The oaks were grown in the field for 4 years, in either current ambient or elevated (current À1 ambient 350 mmol mol ) Ca , in open-top chambers (OTCs). For Q. myrtifolia,an À1 increase in Ca from 360 to 710 mmol mol had no direct effect on R at any time during the year. In April 1999, R in young Q. myrtifolia leaves was significantly higher in elevated CaÐthe only evidence for an indirect effect of growth in elevated Ca. Leaf R was significantly correlated with leaf nitrogen (N)concentration for the sun and shade leaves of both the species of oak. -

Bacterial Production and Respiration
Organic matter production % 0 Dissolved Particulate 5 > Organic Organic Matter Matter Heterotrophic Bacterial Grazing Growth ~1-10% of net organic DOM does not matter What happens to the 90-99% of sink, but can be production is physically exported to organic matter production that does deep sea not get exported as particles? transported Export •Labile DOC turnover over time scales of hours to days. •Semi-labile DOC turnover on time scales of weeks to months. •Refractory DOC cycles over on time scales ranging from decadal to multi- decadal…perhaps longer •So what consumes labile and semi-labile DOC? How much carbon passes through the microbial loop? Phytoplankton Heterotrophic bacteria ?? Dissolved organic Herbivores ?? matter Higher trophic levels Protozoa (zooplankton, fish, etc.) ?? • Very difficult to directly measure the flux of carbon from primary producers into the microbial loop. – The microbial loop is mostly run on labile (recently produced organic matter) - - very low concentrations (nM) turning over rapidly against a high background pool (µM). – Unclear exactly which types of organic compounds support bacterial growth. Bacterial Production •Step 1: Determine how much carbon is consumed by bacteria for production of new biomass. •Bacterial production (BP) is the rate that bacterial biomass is created. It represents the amount of Heterotrophic material that is transformed from a nonliving pool bacteria (DOC) to a living pool (bacterial biomass). •Mathematically P = µB ?? µ = specific growth rate (time-1) B = bacterial biomass (mg C L-1) P= bacterial production (mg C L-1 d-1) Dissolved organic •Note that µ = P/B matter •Thus, P has units of mg C L-1 d-1 Bacterial production provides one measurement of carbon flow into the microbial loop How doe we measure bacterial production? Production (∆ biomass/time) (mg C L-1 d-1) • 3H-thymidine • 3H or 14C-leucine Note: these are NOT direct measures of biomass production (i.e. -
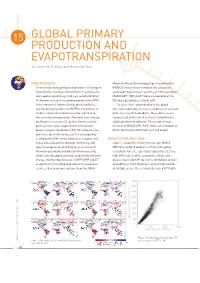
GLOBAL PRIMARY PRODUCTION and EVAPOTRANSPIRATION by Steven W
15 GLOBAL PRIMARY PRODUCTION AND EVAPOTRANSPIRATION by Steven W. Running and Maosheng Zhao DATA PRODUCTS Moderate Resolution Imaging Spectroradiometer Terrestrial primary production provides the energy to (MODIS) sensor, these variables are calculated maintain the structure and functions of ecosystems, every eight days in near real time at 1 km resolution. and supplies goods (e.g. food, fuel, wood and fi bre) MODIS GPP, NPP and ET data are available at the for human society. Gross primary production (GPP) EOS data gateway (see link below). is the amount of carbon fi xed by photosynthesis, To correct for contamination in the global and net primary production (NPP) is the amount of refl ectance data due to severe cloudiness or aerosols carbon converted into biomass after subtracting in the near real time products, these datasets are the cost of plant respiration. The water loss through reprocessed at the end of each year to build more exchange of trace gas CO2 by leaf stomata during stable, permanent datasets. These end-of-year photosynthesis plus evaporation from soil and versions of MODIS GPP, NPP and ET are available at plants is evapotranspiration (ET). ET computes the NTSG, University of Montana (see link below). water lost by a land surface, so it is consequently a component of the water balance in a region, and RESULTS FOR 2000–2006 is therefore relevant for drought monitoring and Figure 1 shows the seven-year average MODIS water management, providing an assessment of NPP for vegetated land on earth at 1 km spatial the water potentially available for human society. -

Phytoplankton As Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate
sustainability Review Phytoplankton as Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate Samarpita Basu * ID and Katherine R. M. Mackey Earth System Science, University of California Irvine, Irvine, CA 92697, USA; [email protected] * Correspondence: [email protected] Received: 7 January 2018; Accepted: 12 March 2018; Published: 19 March 2018 Abstract: The world’s oceans are a major sink for atmospheric carbon dioxide (CO2). The biological carbon pump plays a vital role in the net transfer of CO2 from the atmosphere to the oceans and then to the sediments, subsequently maintaining atmospheric CO2 at significantly lower levels than would be the case if it did not exist. The efficiency of the biological pump is a function of phytoplankton physiology and community structure, which are in turn governed by the physical and chemical conditions of the ocean. However, only a few studies have focused on the importance of phytoplankton community structure to the biological pump. Because global change is expected to influence carbon and nutrient availability, temperature and light (via stratification), an improved understanding of how phytoplankton community size structure will respond in the future is required to gain insight into the biological pump and the ability of the ocean to act as a long-term sink for atmospheric CO2. This review article aims to explore the potential impacts of predicted changes in global temperature and the carbonate system on phytoplankton cell size, species and elemental composition, so as to shed light on the ability of the biological pump to sequester carbon in the future ocean. -

Exchange Velocity Approach and Obt Formation in Plants During the Daytime
EXCHANGE VELOCITY APPROACH AND OBT FORMATION IN PLANTS DURING THE DAYTIME Anca Melintescu PhD “Horia Hulubei” National Institute of Physics and Nuclear Engineering, Bucharest - Magurele, ROMANIA [email protected], [email protected] Third Technical Meeting of the EMRAS II, Working Group 7, “Tritium” Accidents, Vienna, Austria, 24 - 28 January 2011 THE DRIVING EQUATIONS FOR TRITIUM TRANSFER IN ATMOSPHERE - SOIL- PLANT CONTINUUM Driving equation for the HTO transfer from atmosphere to leaves: C – HTO concentration in plant water (Bq/kg); 3 depends on canopy resistance Cair – HTO concentration in air (Bq/m ); Cs - HTO concentration in the sap water (Bq/kg); 3 s - saturated air humidity at vegetation temp. (kg/m ); dC Vexc Vexc - air humidity at reference level (kg/m3); 2 (Cair 0.91 sC) (s )Cs Mw – water mass in plant on a unit soil surface (kg/m ); dt M w M w Vexc – exchange velocity from atmosphere to canopy (m/s) the transpiration flux - used for all canopy, ignoring the transfer of air HTO to steam, because the exchange velocity is smaller with one order of magnitude; - Ignores the initial diffusion of leaf water to steams The tritium dynamics at soil surface: depends on soil resistance dCsw,1 Vex,s C - HTO concentration in the first soil layer at the (Bq/kg); (Cair 0.91sat (Ts )Csw,1 ) DF sw,1 Vex,s - exchange velocity from atmosphere to soil (m/s); dt M 3 ws sat(Ts) - saturated air humidity at soil surface temp. (kg/m ); Mws – water mass in the surface soil layer; DF - HTO net flux at the bottom interface of the first soil -

Drought and Ecosystem Carbon Cycling
Agricultural and Forest Meteorology 151 (2011) 765–773 Contents lists available at ScienceDirect Agricultural and Forest Meteorology journal homepage: www.elsevier.com/locate/agrformet Review Drought and ecosystem carbon cycling M.K. van der Molen a,b,∗, A.J. Dolman a, P. Ciais c, T. Eglin c, N. Gobron d, B.E. Law e, P. Meir f, W. Peters b, O.L. Phillips g, M. Reichstein h, T. Chen a, S.C. Dekker i, M. Doubková j, M.A. Friedl k, M. Jung h, B.J.J.M. van den Hurk l, R.A.M. de Jeu a, B. Kruijt m, T. Ohta n, K.T. Rebel i, S. Plummer o, S.I. Seneviratne p, S. Sitch g, A.J. Teuling p,r, G.R. van der Werf a, G. Wang a a Department of Hydrology and Geo-Environmental Sciences, Faculty of Earth and Life Sciences, VU-University Amsterdam, De Boelelaan 1085, 1081 HV Amsterdam, The Netherlands b Meteorology and Air Quality Group, Wageningen University and Research Centre, P.O. box 47, 6700 AA Wageningen, The Netherlands c LSCE CEA-CNRS-UVSQ, Orme des Merisiers, F-91191 Gif-sur-Yvette, France d Institute for Environment and Sustainability, EC Joint Research Centre, TP 272, 2749 via E. Fermi, I-21027 Ispra, VA, Italy e College of Forestry, Oregon State University, Corvallis, OR 97331-5752 USA f School of Geosciences, University of Edinburgh, EH8 9XP Edinburgh, UK g School of Geography, University of Leeds, Leeds LS2 9JT, UK h Max Planck Institute for Biogeochemistry, PO Box 100164, D-07701 Jena, Germany i Department of Environmental Sciences, Copernicus Institute of Sustainable Development, Utrecht University, PO Box 80115, 3508 TC Utrecht, The Netherlands j Institute of Photogrammetry and Remote Sensing, Vienna University of Technology, Gusshausstraße 27-29, 1040 Vienna, Austria k Geography and Environment, Boston University, 675 Commonwealth Avenue, Boston, MA 02215, USA l Department of Global Climate, Royal Netherlands Meteorological Institute, P.O. -

Detailed Temporal Modelling of Carbon and Water Fluxes from Pastures in New Zealand
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere without the permission of the Author. Detailed temporal modelling of carbon and water fluxes from pastures in New Zealand: Case study of an experimental dairy farm in the Waikato region A thesis presented in partial fulfilment of the Requirements for the degree of Doctor of Philosophy In Soil Science Massey University, Palmerston North, New Zealand Nicolas Puche 2017 i ii ABSTRACT The terrestrial biosphere is an important pool of carbon, with its size governed by the opposing processes of CO2 uptake through photosynthesis and release through respiration. It is therefore critically important to understand and reliably and accurately model these processes and predict changes in carbon exchange in response to key drivers. Pasture-based livestock production is particularly important for the New Zealand’s economy but it is also a main contributor to NZ’s greenhouse gas budget. My Ph.D. work used half-hourly eddy-covariance (EC) data, previously collected over 2 consecutive years from a grazed pasture in the Waikato region. The main aims of this study were to assess whether there was any bias in gap-filled eddy covariance measurements, to assess whether incomplete capture of cow respiration during grazing events could have led to biased observations, and to quantify the resulting difference on the net carbon budget of the farm. I approached the work by developing a new process-based model, CenW_HH, running at a half-hourly time step, to predict the energy and CO2 exchange of grazed pastures. -

Grassland to Shrubland State Transitions Enhance Carbon Sequestration in the Northern Chihuahuan Desert
Global Change Biology Global Change Biology (2015) 21, 1226–1235, doi: 10.1111/gcb.12743 Grassland to shrubland state transitions enhance carbon sequestration in the northern Chihuahuan Desert M. D. PETRIE1 ,S.L.COLLINS1 ,A.M.SWANN2 ,P.L.FORD3 andM.E. LITVAK1 1Department of Biology, University of New Mexico, Albuquerque, New Mexico, USA, 2Department of Biology, Sevilleta LTER, University of New Mexico, Albuquerque, New Mexico, USA, 3USDA Forest Service, Rocky Mountain Research Station, Albuquerque, New Mexico, USA Abstract The replacement of native C4-dominated grassland by C3-dominated shrubland is considered an ecological state tran- sition where different ecological communities can exist under similar environmental conditions. These state transi- tions are occurring globally, and may be exacerbated by climate change. One consequence of the global increase in woody vegetation may be enhanced ecosystem carbon sequestration, although the responses of arid and semiarid ecosystems may be highly variable. During a drier than average period from 2007 to 2011 in the northern Chihuahuan À2 À1 Desert, we found established shrubland to sequester 49 g C m yr on average, while nearby native C4 grassland À À was a net source of 31 g C m 2 yr 1 over this same period. Differences in C exchange between these ecosystems were pronounced – grassland had similar productivity compared to shrubland but experienced higher C efflux via ecosys- tem respiration, while shrubland was a consistent C sink because of a longer growing season and lower ecosystem respiration. At daily timescales, rates of carbon exchange were more sensitive to soil moisture variation in grassland than shrubland, such that grassland had a net uptake of C when wet but lost C when dry. -

Model-Based Analysis of the Impact of Diffuse
Agricultural and Forest Meteorology 249 (2018) 377–389 Contents lists available at ScienceDirect Agricultural and Forest Meteorology journal homepage: www.elsevier.com/locate/agrformet Model-based analysis of the impact of diffuse radiation on CO2 exchange in a T temperate deciduous forest Min S. Leea, David Y. Hollingerb, Trevor F. Keenanc, Andrew P. Ouimetted, Scott V. Ollingerd, ⁎ Andrew D. Richardsona,e,f, a Harvard University, Department of Organismic and Evolutionary Biology, 26 Oxford Street, Cambridge, MA 02138, USA b USDA Forest Service, Northern Research Station, 271 Mast Rd, Durham, NH 03824, USA c Climate and Ecosystem Sciences Division, Lawrence Berkeley National Lab., 1 Cyclotron Rd., Berkeley, CA 94720, USA d University of New Hampshire, Earth Systems Research Center, 8 College Rd, Durham, NH 03824, USA e Northern Arizona University, School of Informatics, Computing and Cyber Systems, PO Box 5693, Flagstaff, AZ 86011, USA f Northern Arizona University, Center for Ecosystem Science and Society, PO Box 5620, Flagstaff, AZ 86011, USA ARTICLE INFO ABSTRACT Keywords: Clouds and aerosols increase the fraction of global solar irradiance that is diffuse light. This phenomenon is Diffuse radiation known to increase the photosynthetic light use efficiency (LUE) of closed-canopy vegetation by redistributing Light use efficiency photosynthetic photon flux density (400–700 nm) from saturated, sunlit leaves at the top of the canopy, to Eddy covariance shaded leaves deeper in the canopy. We combined a process-based carbon cycle model with 10 years of eddy Net ecosystem exchange covariance carbon flux measurements and other ancillary data sets to assess 1) how this LUE enhancement Deciduous forest influences interannual variation in carbon uptake, and 2) how errors in modeling diffuse fraction affect pre- Canopy photosynthesis dictions of carbon uptake. -

Respiration from a Tropical Forest Ecosystem: Partitioning of Sources and Low Carbon Use Efficiency
Ecological Applications, 14(4) Supplement, 2004, pp. S72±S88 q 2004 by the Ecological Society of America RESPIRATION FROM A TROPICAL FOREST ECOSYSTEM: PARTITIONING OF SOURCES AND LOW CARBON USE EFFICIENCY JEFFREY Q. CHAMBERS,1,2,4 EDGARD S. TRIBUZY,2 LIGIA C. TOLEDO,2 BIANCA F. C RISPIM,2 NIRO HIGUCHI,2 JOAQUIM DOS SANTOS,2 ALESSANDRO C. ARAUÂ JO,2 BART KRUIJT,3 ANTONIO D. NOBRE,2 AND SUSAN E. TRUMBORE1 1University of California, Earth System Sciences, Irvine, California 92697 USA 2Instituto Nacional de Pesquisas da AmazoÃnia, C.P. 478, Manaus, Amazonas, Brazil, 69011-970 3Alterra, University of Wageningen, Alterra, 6700 AA Wageningen, The Netherlands Abstract. Understanding how tropical forest carbon balance will respond to global change requires knowledge of individual heterotrophic and autotrophic respiratory sources, together with factors that control respiratory variability. We measured leaf, live wood, and soil respiration, along with additional environmental factors over a 1-yr period in a Central Amazon terra ®rme forest. Scaling these ¯uxes to the ecosystem, and combining our data with results from other studies, we estimated an average total ecosystem respiration (Reco) of 7.8 mmol´m22´s21. Average estimates (per unit ground area) for leaf, wood, soil, total heterotrophic, and total autotrophic respiration were 2.6, 1.1, 3.2, 5.6, and 2.2 mmol´m22´s21, respectively. Comparing autotrophic respiration with net primary production (NPP) esti- mates indicated that only ;30% of carbon assimilated in photosynthesis was used to con- struct new tissues, with the remaining 70% being respired back to the atmosphere as autotrophic respiration.